Прионные болезни человека
Что же касается средств иммунотерапии и, естественно, иммунопрофилактики, то здесь пока не существует реальных оснований, которые позволяли бы рассчитывать на успех по крайней мере в обозримом будущем, в связи с тем что инфекционный прионный белок PrPSc иммунологически не отличим от нормального прионного белка PrPC.
Вторая особенность прионных болезней обусловлена тем, что они представляют собой неотъемлемую часть теперь уже достаточно обширной (около 40 нозологических форм) группы медленных инфекций человека и животных. Как известно, подавляющее большинство этих заболеваний вызывают вирусы, известные как возбудители острых инфекций. Это лишний раз подчеркивает справедливость утверждения о том, что большинство вирусов в зависимости от условий заражения (или пребывания) способствует развитию в организме различных форм инфекционного процесса.
В связи с этим прионные болезни занимают особое положение, так как их возбудители не способны к столь выраженной универсальности, как у вирусов, и они (инфекционные прионные белки - PrPSc) не формируют и не поддерживают в организме иные процессы, кроме медленного и (как это было установлено уже давно и впоследствии неоднократно подтверждалось экспериментально) бессимптомного.
Отмеченная особенность, т.е. неспособность вызывать острую форму инфекционного процесса, по-видимому, обусловлена особенностями самих возбудителей прионных болезней, так как уже давно обнаружено, что сам процесс накопления инфекционного прионного белка PrPSc в различных органах и тканях экспериментально зараженного лабораторного животного протекает весьма медленно. Можно полагать, что низкая скорость накопления инфекционного агента в данном случае обусловлена событиями, лежащими в основе механизма превращения клеточного прионного белка (PrPC) в инфекционный прионный белок (PrPSc).
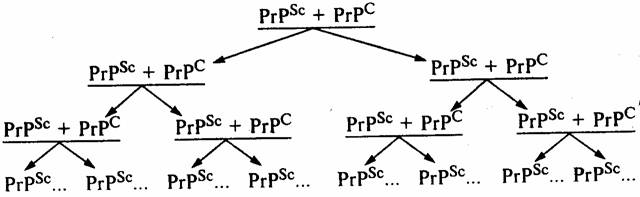
Собственно механизм накопления инфекционного белка в зараженном организме сегодня точно неизвестен. Вместе с тем, исходя из определения, что это посттрансляционный процесс, очевидно, что инфекционный прионный белок вызывает в здоровом организме трансформацию нормального прионного белка в инфекционную форму за счет его (нормального белка) конформационных (т.е. пространственных) изменений. В этом случае речь идет об изменении третичной или даже четвертичной структуры исходного белка PrPC. Таким образом, процесс накопления инфекционного прионного белка происходит не в результате синтеза в зараженном организме молекул PrPSc de novo, а вследствие конформационных изменений уже синтезированных перед этим нормальных молекул PrPC под влиянием инфекционного прионного белка PrPSc (схема). Процесс накопления инфекционного прионного белка обусловлен прежде всего необходимостью контакта двух молекул. В результате под влиянием одной молекулы PrPSc происходит трансформация одной молекулы PrPC в ее инфекционную форму PrPSc. Следующий этап, как видно на схеме, включает в себя уже наличие влияния двух молекул PrPSc, под воздействием которых образуются уже четыре молекулы PrPSc и т.д. Таким образом, как видно из приведенной схемы, процесс накопления инфекционного прионного белка носит лавинообразный характер.
Процесс накопления молекул инфекционного прионного белка
СТРУКТУРА ПРИОННЫХ БЕЛКОВ
Установленные необычные свойства возбудителей ТГЭ послужили основанием для выдвижения большого количества разнообразных теорий, пытающихся объяснить структуру и химическую природу этих агентов, многие из которых теперь имеют лишь историческое значение. Резкий скачок вперед в понимании природы возбудителей ТГЭ был сделан в результате разработок эффективных методов очистки и концентрации агента скрепи. Существенный вклад в разработку таких методов внесла группа Стенли Прузинера из Калифорнийского университета (США). Разработанная им многоступенчатая система очистки позволила получить препараты, очищенные в 100 – 1000 раз. На основании изучения высокоочищенных препаратов авторы пришли к выводу о том, что возбудитель скрепи является белком. Этот вывод был сделан в результате анализа инактивации агента при его обработке протеазой К, модификации при воздействии диэтилпирокарбонатом, додецилсульфатом натрия, гуанидинтиоцианатом, фенолом и мочевиной. Агент оставался устойчивым к обработке рядом реагентов, инактивирующих нуклеиновые кислоты, что указывало на их отсутствие в его составе. Изучение очищенного препарата возбудителя скрепи показало, что он обладает молекулярной массой около или меньше 50 000 Да.
Следует отметить, что представление о прионной природе возбудителя скрепи, выдвинутое С.Прузинером, оказалось очень плодотворным и послужило основанием для более детального распознавания природы возбудителей ТГЭ. В результате дальнейшей очистки приона было показано, что его основным компонентом является мажорный белок с молекулярной массой 27000 – 30000 Да, обозначаемый как РrР 27–30. Этот белок является составной частью скрепи-ассоциированных фибрилл, причем получены структурные и биохимические свидетельства того, что сборка этих фибрилл происходит in vivo, и изучены некоторые молекулярные механизмы их образования. По своей физико-химической характеристике РrР представляет собой сиалогликопротеин и является первым идентифицированным структурным компонентом приона скрепи. Появление РrР 27–30 на этапе развития инфекции до развития гистопатологических изменений указывало на то, что этот белок не является вторичным продуктом патологической реакции. Был сделан вывод о том, что РrР 27–30 играет центральную роль в патогенезе скрепи.
При дальнейшем изучении прионов, выделенных из головного мозга зараженных скрепи животных, было выявлено наличие в ЦНС частиц в виде стержней диаметром 10 – 20 нм и длиной 100 – 200 нм. Ультраструктурно они напоминали амилоид и, по-видимому, представляли собой полимерную форму приона скрепи; каждый стержень содержал около 1000 молекул приона. Был проанализирован аминокислотный состав PrP 27–30 и определена последовательность 15 аминокислотных остатков в его полипептидной цепи. В последующем из головного мозга зараженных скрепи хомяков был выделен мажорный белок с молекулярной массой 33–37 кДа, обозначенный как HaSp 33–37; его выделение проводилось без этапа обработки протеазами. Обработка HaSp 33–37 протеазой К приводила к получению продукта, электрофоретически неотличимого от РrР 27–30. Была определена последовательность 22 аминокислотных остатков HaSp 33–37. Авторы полагали, что HaSp 33–37 представляет собой интактную форму белка возбудителя скрепи. Были изучены также некоторые другие характеристики прионов скрепи и болезни Крейтцфельдта–Якоба. В частности, при изучении липосом было подтверждено предположение о том, что инфекционная частица скрепи содержит 2 молекулы PrPSc и показано наличие вставок в ген приона при семейных случаях болезни Крейтцфельдта–Якоба и синдрома Герстманна–Штреусслера–Шейнкера.
Важным шагом, имеющим как теоретическое, так и методическое значение, было получение антител при использовании в качестве антигенов высокоочищенных прионов скрепи. В сыворотках кроликов, иммунизированных РrР 27–30, определяли антитела, специфически реагирующие с РrР 27–30 и с несколькими белками с более низкой молекулярной массой, очевидно, имеющими общую антигенную детерминанту с РrР 27–30 или являющимися продуктами его расщепления. Полученные антисыворотки не взаимодействовали с соответствующими белками, выделенными из головного мозга нормальных незараженных животных. Используя полученную антисыворотку с пероксидазной меткой, удалось показать локализацию прионов в определенных отделах головного мозга зараженных животных. В соответствии с ранее полученными данными структуры, связанные с меченой антисывороткой, обладали характеристикой амилоидных бляшек. Получение и использование антисыворотки к синтетическому пептиду, соответствующему N-концевой части приона скрепи, позволили провести индикацию белка скрепи-ассоциированных фибрилл в головном мозге, селезенке и лимфатических узлах зараженных животных. При этом положительные результаты были получены на ранних этапах инкубационного периода скрепи.