Отбор в менделевской бесполой диплоидной популяцииРефераты >> Математика >> Отбор в менделевской бесполой диплоидной популяции
Закон Харди -Вайнберга гласит, что при отсутствии возмущений частоты аллелей не изменяются. Однако процессы, изменяющие частоты генов, постоянно происходят в популяции. Без них не было бы эволюции. Рассмотрим процессы отбора. К идее естественного отбора как основного процесса в эволюции пришли независимо друг от друга Чарльз Дарвин и Альфред Рассел Уоллес. В 1858 г. на заседании Линнеевского общества в Лондоне были представлены сообщения об их открытии. Доказательства того, что эволюция происходит путем естественного отбора были даны Ч. Дарвином в его работе "Происхождение видов" в 1859 г. Ч. Дарвин предположил, что в результате изменений появляются наследуемые признаки, которые обеспечивают их обладателям лучшую приспособленность к условиям среды. Такие организмы имеют больше шансов к выживанию и оставляют больше потомства, чем особи наделенные менее полезными свойствами. В результате частота признаков, обеспечивающих лучшую приспособленность (адаптивность), будет увеличиваться от поколения к поколению за счет частоты признаков, дающих меньшую адаптивность. Этот процесс был назван естественным отбором.
Рассмотрим простейший случай - модель отбора в двухаллельной менделевской популяции. В ее основе лежит закон Харди -Вайнберга расчета частот аллелей и генотипов. Припишем генотипам AA, Aa, aa числа ![]() ,
, ![]() ,
, ![]() , которые назовем относительными приспособленностями. Данные коэффициенты можно интерпретировать как вероятности того, что особь соответствующего генотипа доживает до этапа размножения. Обозначим через
, которые назовем относительными приспособленностями. Данные коэффициенты можно интерпретировать как вероятности того, что особь соответствующего генотипа доживает до этапа размножения. Обозначим через ![]() и
и ![]() частоты
частоты ![]() аллелей A и a в n -ом поколении в момент его появления (они же - частоты гамет n-1 ого поколения). Первоначально для n -ого поколения согласно закону Харди -Вайнберга для генотипов AA, Aa, aa имеем частоты:
аллелей A и a в n -ом поколении в момент его появления (они же - частоты гамет n-1 ого поколения). Первоначально для n -ого поколения согласно закону Харди -Вайнберга для генотипов AA, Aa, aa имеем частоты: ![]() ,
, ![]() ,
, ![]() . Числа
. Числа ![]() ,
, ![]() ,
, ![]() есть вероятности того, что особь n -ого поколения относится соответственно к генотипу AA, Aa, aa и доживает до этапа размножения. Полная вероятность того, что особь n - ого поколения доживает до размножения суть
есть вероятности того, что особь n -ого поколения относится соответственно к генотипу AA, Aa, aa и доживает до этапа размножения. Полная вероятность того, что особь n - ого поколения доживает до размножения суть
![]() . (13)
. (13)
Эту величину будем называть средней приспособленностью в n -ом поколении. Используя формулу Бейеса (теорема гипотез), выписываем частоты генотипов AA, Aa, aa этого поколения к моменту начала этапа размножения:
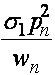 ,
, 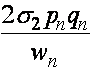 ,
, 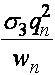 .
.
Частоты гамет A, a определяются частотами генотипов:
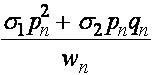 ,
, 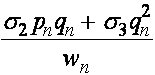 .
.
Частоты аллелей A, a в момент появления n+1 -ого поколения совпадают с частотами гамет предыдущего поколения. Получаем уравнения связывающие частоты аллелей:
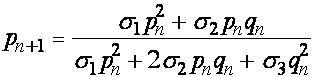 , (14)
, (14)
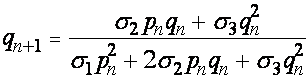 . (15)
. (15)
Независимым из этих двух уравнений является только одно т.к. ![]() . Отметим, что уравнения (14) и (15) не меняются, если заменить
. Отметим, что уравнения (14) и (15) не меняются, если заменить ![]() , где
, где ![]() произвольно. В связи с этим часто принимают равной единице максимальную относительную приспособленность.
произвольно. В связи с этим часто принимают равной единице максимальную относительную приспособленность.
Уравнение (14) имеет явные решения только в частных случаях. Рассмотрим случай так называемого геометрического отбора. Пусть относительные приспособленности генотипов образуют геометрическую прогрессию: ![]() ,
, ![]() ,
, ![]() . Введем обозначения
. Введем обозначения 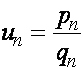 . Разделив первое уравнение на второе, получим
. Разделив первое уравнение на второе, получим
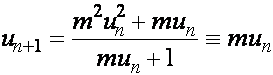 .
.
Отсюда следует, что ![]() . Вернемся к старым переменным:
. Вернемся к старым переменным:
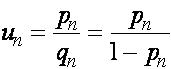 .
.
Получаем
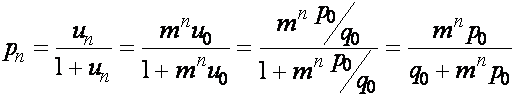 .
.
Если ![]() , то отбора нет,
, то отбора нет, ![]() и для всех
и для всех ![]() . Если
. Если ![]() , то
, то ![]() и
и ![]() с ростом
с ростом ![]() , т.е. аллель A вытесняет аллель a. Если
, т.е. аллель A вытесняет аллель a. Если ![]() , то наоборот -аллель a вытесняет аллель A. Отбор геометрического типа эквивалентен отбору на уровне аллелей. Действительно, пусть
, то наоборот -аллель a вытесняет аллель A. Отбор геометрического типа эквивалентен отбору на уровне аллелей. Действительно, пусть ![]() - вероятность того что, особь, имеющая один аллель A, доживет до стадии размножения. В свою очередь, пусть
- вероятность того что, особь, имеющая один аллель A, доживет до стадии размножения. В свою очередь, пусть ![]() аналогичная вероятность по аллелю a. Данные вероятности можно понимать как вероятности выживания аллелей и можно назвать приспособленностью аллелей. Если считать, что гибель аллеля приводит к гибели особи, то вероятности выживания (приспособленности) особей генотипов AA, Aa, aa суть
аналогичная вероятность по аллелю a. Данные вероятности можно понимать как вероятности выживания аллелей и можно назвать приспособленностью аллелей. Если считать, что гибель аллеля приводит к гибели особи, то вероятности выживания (приспособленности) особей генотипов AA, Aa, aa суть ![]() ,
, ![]() ,
, ![]() . Как уже отмечалось, приспособленности можно нормировать (разделить на одно и то же число). После деления на
. Как уже отмечалось, приспособленности можно нормировать (разделить на одно и то же число). После деления на ![]() получим приспособленности генотипов
получим приспособленности генотипов ![]() ,
, ![]() ,
, ![]() , где
, где 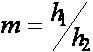 .
.