МиграцииРефераты >> Математика >> Миграции
Миграции, или поток генов возникают, когда особи одной популяции перемещаются в другую и скрещиваются с членами второй популяции. Поток генов не меняют частоты аллелей у вида в целом. Однако, частоты могут меняться в локальных популяциях, если исходные частоты различны у старожилов и пришельцев.
Рассмотрим простейшую модель, описывающую локальную популяцию, в которую с определенной частотой мигрируют особи из окружающей популяции и скрещиваются со старожилами. Предположим, что частота аллеля ![]() в окружающей популяции постоянна и равна
в окружающей популяции постоянна и равна ![]() . Пусть
. Пусть ![]() -вероятность обнаружения пришельца среди особей текущего поколения (доля мигрантов). Эта вероятность характеризует интенсивность миграции. Считаем, что доля пришельцев для всех поколений одна и та же.
-вероятность обнаружения пришельца среди особей текущего поколения (доля мигрантов). Эта вероятность характеризует интенсивность миграции. Считаем, что доля пришельцев для всех поколений одна и та же.
Обозначим частоту аллеля ![]() для локальной популяции через
для локальной популяции через ![]() . Тогда частота для следующего поколения суть:
. Тогда частота для следующего поколения суть:
![]() .
.
Здесь ![]() -вероятность того, что гамета
-вероятность того, что гамета ![]() -ого поколения является старожилом и обладает аллелем
-ого поколения является старожилом и обладает аллелем ![]() . Соответственно,
. Соответственно, ![]() -вероятность того, что гамета является пришельцем и имеет аллель
-вероятность того, что гамета является пришельцем и имеет аллель ![]() . Положим:
. Положим: ![]() . В результате получим:
. В результате получим:
![]() ,
,
![]() .
.
Поскольку ![]() , то то с ростом номера поколения
, то то с ростом номера поколения ![]() и
и ![]() . Частота аллеля
. Частота аллеля ![]() в локальной популяции уравниваетсяс его частотой во внешней популяции. Полученную формулу можно использовать для оценки интенсивности потока генов. Рассмотрим пример.
в локальной популяции уравниваетсяс его частотой во внешней популяции. Полученную формулу можно использовать для оценки интенсивности потока генов. Рассмотрим пример.
В США потомство от смешанных браков между белыми и темнокожими принято относить к темнокожему населению. Следовательно, смешанные браки можно рассматривать как поток генов из белой в темнокожую популяцию. Частота аллеля ![]() , контролирующего резус фактор, у белого населения США составляет
, контролирующего резус фактор, у белого населения США составляет ![]() . В африканских племенах, от которых происходит современное темнокожее население США, частота этого аллеля
. В африканских племенах, от которых происходит современное темнокожее население США, частота этого аллеля ![]() . Предки современных темнокожих США были вывезены из Африки примерно
. Предки современных темнокожих США были вывезены из Африки примерно ![]() лет назад (около
лет назад (около ![]() поколений), следовательно на данный момент
поколений), следовательно на данный момент ![]() . Частота аллеля
. Частота аллеля ![]() у современного темнокожего населения США
у современного темнокожего населения США ![]() . Получаем:
. Получаем:
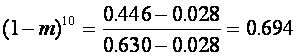 .
.
![]() ,
, ![]() .
.
Число ![]() -вероятность того, что данный аллель не являлся мигрантом ни в одном из десяти предшествующих поколений. Тем самым, современное темнокожее население США наследовало около
-вероятность того, что данный аллель не являлся мигрантом ни в одном из десяти предшествующих поколений. Тем самым, современное темнокожее население США наследовало около ![]() генов от своих африканских предков, а
генов от своих африканских предков, а ![]() генов получило от белых предков. Неудивительно, что темнокожие американцы по внешнему виду существенно отличаются от африканцев. Отметим, что число
генов получило от белых предков. Неудивительно, что темнокожие американцы по внешнему виду существенно отличаются от африканцев. Отметим, что число ![]() примерно соответствует проценту смешанных браков.
примерно соответствует проценту смешанных браков.
Приведенные вкладки носят приближенный характер. Если в аналогичных расчетах использовать частоты других аллелей, то получатся несколько другие результаты. Кроме того, интенсивность потока генов в разных регионах США различна. Однако, тенденция к выравниванию частот аллелей между темнокожим и белым населением сохраняется.
Рассмотрим островную модель Райта. Эта модель учитывает как миграции, так и процессы отбора. Она описывает популяцию аллелей ![]() и
и ![]() , которая состоит из
, которая состоит из ![]() субпопуляций. Обозначим через
субпопуляций. Обозначим через ![]() , где
, где ![]() , доли субпопуляций (вероятности того, что что выбранный аллель принадлежит
, доли субпопуляций (вероятности того, что что выбранный аллель принадлежит ![]() -ой популяции). Будем считать, что эти доли из поколения в поколение неизменны. Субпопуляции связаны между собой меграционными процессами. Непосредственно после появления на свет нового поколения из каждой субполяции с вероятностью
-ой популяции). Будем считать, что эти доли из поколения в поколение неизменны. Субпопуляции связаны между собой меграционными процессами. Непосредственно после появления на свет нового поколения из каждой субполяции с вероятностью ![]() аллели (их носители) совершают путешествие в некий “общий котел” аллелей (внешний мир). Считаем, что вероятность
аллели (их носители) совершают путешествие в некий “общий котел” аллелей (внешний мир). Считаем, что вероятность ![]() миграции для всех субпопуляций одна и та же. В “общем котле” аллели перемешиваются, “забывают” о своем “географическом” происхождении и мигрируют в субпопуляции. Пусть вероятность обнаружить вернувшихся путешественников (доля мигрантов) для всех субпопуляций одна и та же и равна
миграции для всех субпопуляций одна и та же. В “общем котле” аллели перемешиваются, “забывают” о своем “географическом” происхождении и мигрируют в субпопуляции. Пусть вероятность обнаружить вернувшихся путешественников (доля мигрантов) для всех субпопуляций одна и та же и равна ![]() , т.е. совпадает с вероятностью исхода юных аллелй из субпопуляций. Далее, в субпопуляциях начинается процесс отбора. Он происходит на уровне аллелей (для генотипов имеет место отбор геометрического типа). Пусть
, т.е. совпадает с вероятностью исхода юных аллелй из субпопуляций. Далее, в субпопуляциях начинается процесс отбора. Он происходит на уровне аллелей (для генотипов имеет место отбор геометрического типа). Пусть ![]() и
и ![]() -вероятности того, что в
-вероятности того, что в ![]() -ой субпопуляции соответственно аллели
-ой субпопуляции соответственно аллели ![]() и
и ![]() доживают до этапа размножения (эти числа также называются коэффициентами отбора).
доживают до этапа размножения (эти числа также называются коэффициентами отбора).