Концепция материальности сознанияРефераты >> Естествознание >> Концепция материальности сознания
Синапсы выполняют функцию усилителей нервных сигналов на пути их следования. Эффект достигается тем, что один относительно маломощный электрический импульс освобождает сотни тысяч молекул медиатора, заключенных до того во многих синаптических пузырьках. Залп молекул медиатора синхронно действует на небольшой участок управляемого нейрона, где сосредоточены постсинаптические рецепторы - специализированные белки, которые преобразуют сигнал теперь уже из химической формы в электрическую.
Синапсы - удобный объект регулирования потоков информации. Уровень усиления сигнала при его передаче через синапс можно легко увеличить или уменьшить, изменяя количество освобождаемого медиатора, вплоть до полного запрета на передачу информации. Теоретически это можно осуществить путем направленного воздействия на любой из этапов высвобождения медиатора.
Именно на выяснение механизмов управления синаптической передачей и были устремлены усилия исследователей в лаборатории биофизики Института эволюционной физиологии и биохимии им. И. М. Сеченова РАН (Санкт-Петербург). Их эффективному проведению способствовала поддержка Международного научного фонда, позволив, в частности, оперативно приобрести современные приборы и разнообразные химические реактивы.
В качестве подходящего объекта исследования выбрали мышцу лягушки, изолированную вместе с управляющим нервом, такой препарат обладает главными достоинствами: однотипностью синапсов, удобством проведения разнообразных и достаточно тонких экспериментов, способностью изолированных тканей длительное время сохранять свою жизнеспособность. Поскольку основные свойства синапсов в разных тканях весьма сходны, полученные сведения можно использовать для анализа синаптической передачи в мозгу.
Свойства нервно-мышечных синапсов диктовали и методы регистрации. Одиночный акт передачи сигнала через синапс длится всего несколько тысячных долей секунды (миллисекунд), поэтому для его регистрации возможны только электрические измерения. С этой целью изолированный препарат помещают в ванночку с солевым раствором, с той же концентрацией ионов, как в плазме крови лягушки. Нерв укладывают на раздражающие электроды, а в мышечное волокно, в непосредственной близости с синапсом, вводят внутриклеточные микроэлектроды. Последние представляют собой тонкие стеклянные трубки, диаметр вытянутого кончика которых не превышает одного-двух микрометров. Микроэлектрод, заполненный концентрированным раствором хлористого калия, может служить для измерения разности потенциалов между внутренностью мышечного волокна и его поверхностью. Специальная система усилителей биоэлектрических сигналов позволяет одновременно регистрировать и быстрые изменения электрического тока, вызванные действием медиатора, и удерживать потенциал на мембране мышечного волокна на заданном уровне. Электрические синаптические ответы затем можно вывести на экран осциллографа (рис.4), записать на магнитную ленту либо после преобразования в цифровую форму проанализировать на компьютере. Такая система позволяет экспериментатору оценивать изменения синаптической передачи в ходе опыта.
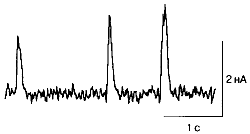 Спонтанные постсинаптические токи, генерируемые мышечным волокном. Каждый сигнал - ответ на действие порции медиатора, освободившегося из синаптических пузырьков
Спонтанные постсинаптические токи, генерируемые мышечным волокном. Каждый сигнал - ответ на действие порции медиатора, освободившегося из синаптических пузырьков
|
Прежде всего, какие же химические агенты модулируют освобождение медиатора? Наибольший интерес для нас представляло пресинаптическое действие самих медиаторов. Биологический смысл использования одних медиаторов для регуляции освобождения других очевиден. Такой механизм обеспечивает вмешательство третьего нейрона в "разговор" первых двух: выделяемый им медиатор действует на нервное окончание управляющего нейрона и регулирует освобождение медиатора, предназначенного для передачи сигнала управляемому нейрону. Здесь, как правило, используются медиаторы разной химической природы: один для передачи по основному пути, а другой - для ее модуляции. Но медиатор может и сам управлять своим освобождением. Если из нервного окончания освобождается избыточное количество медиатора, это в конце концов приводит к нерациональным энергетическим затратам на синтез необходимых веществ и на восстановление режима готовности к новому рабочему акту. Наконец, перевозбуждение может стать губительным для управляемой клетки. Значит, необходима саморегулирующаяся система, основанная на принципе отрицательной обратной связи. Действительно, медиатор в нервно-мышечных синапсах, ацетилхолин, будучи выброшен в синаптическую щель, не только выполняет свою прямую обязанность - передать информацию (например, двигательный приказ мышечному волокну, действуя на его постсинаптические рецепторы), но и способен регулировать собственное освобождение, действуя через пресинаптические рецепторы нервных окончаний.
Принципиальная схема регуляции должна выглядеть следующим образом: медиатор действует на пресинаптический рецептор, изменения состояния рецептора передаются через некие промежуточные стадии на системы, способные влиять на выброс медиатора, - и в результате снижается вероятность этого события.
Нужно было ответить на целый ряд вопросов, логически связанных друг с другом. Прежде всего определить, какой тип пресинаптического рецептора вовлечен в процесс саморегулирования. Оказывается, природа и здесь подошла к решению проблемы рационально. Для передачи двигательных сигналов используется один тип рецептора, рассчитанный на быстрое их преобразование, а для регулирования - рецепторы другого типа, работающие гораздо медленнее, но зато обеспечивающие высокий коэффициент усиления сигнала. Несмотря на то, что оба типа рецептора реагируют на один и тот же медиатор (ацетилхолин), в эксперименте, используя различия в их чувствительности к синтезированным химическим аналогам ацетилхолина, можно избирательно действовать на постсинаптические (пусковые) или на пресинаптические (регулирующие) рецепторы (рис.5).
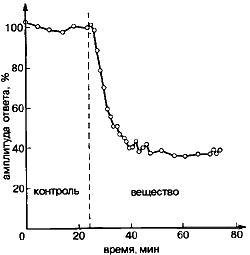 Рис.5.
Рис.5.