Размножение папоротников в оранжерейных условияхРефераты >> Ботаника и сельское хоз-во >> Размножение папоротников в оранжерейных условиях
При выращивании папоротников из спор мы руководствовались методическими рекомендациями ряда авторов (Парамонова, 1956; Чичагова, 1958; Котухов, 1967; Арнаутова, 1987; Мак-Миллан Броуз, 1987; Хессайон, 1995). Для сбора спор срезали фертильную часть листа папоротника и помещали его в пакет из кальки. В течение нескольких дней пакет держали в теплом месте до раксрытия спорангиев и высыпания спор. Споры тонким слоем высевали на земляную смесь в чашки Петри. Земляную смесь готовили из 2 частей простерилизованной листовой земли и 1 части промытого речного песка. Закрытые чашки Петри ставили в затененное место. В чашках постоянно поддерживали высокий уровень влажности. После появления гаметофитов почву время от времени обильно поливали, что необходимо для успешного протекания полового процесса. После окончания полового процесса наблюдается появление проростков спорофита. Для закаливания и адаптации растений к более засушливым условиям в этот период стекло с чашки Петри ежедневно на некоторое время убирали. После появления 3-4-го листа у спорофита проводили пикировку растений. Горшки с распикированными растениями накрывали стеклом. Первые дни растения только опрыскивали, снимая стекло на 2-3 ч в день. После укоренения стекло снимали и полив проводили по мере необходимости.
Для изучения начальных этапов онтогенеза Циртомиума серповидного споры высевали в чашки Петри на жидкую среду Кнопа. Для этого в чашку Петри помещали фильтровальную бумагу, увлажненную раствором Кнопа следующего состава: Ca(NО3)2 - 1 г, MgSO4- 0,25 г, KСl - 0,125 г, KH2PO4 - 0,25 г на 1 л воды.
Собранные споры тонким слоем распределяли на фильтровальной бумаге. Наблюдения за прорастанием спор, формированием гаметофитов и проростков проводили с помощью стереоскопического микроскопа МБС-9 и микроскопа “Биолам”. Наблюдения проводились ежедневно. Полученные данные фиксировались в виде рисунков.
При изучении развития гаметофита Циртомиума мы, следуя другим авторам (Nayar, Kaur, 1971; Арнаутова, 1988, 1989; Храпко, 1990а, 1990б, 1990в, 1991а, 1991б, 1996; Шорина, Ершова, 1990 и др.), выделяли 3 этапа: 1 - прорастание споры; 2 - образование нити гаметофита, 3 - формирование пластинки гаметофита. При изучении развития спорофита Циртомиума серповидного мы придерживались периодизации онтогенеза спорофитов папоротников, предложенной Г.П. Серой с соавторами (1980). В онтогенезе спорофита они выделяют 3 периода - препродуктивный, репродуктивный и пострепродуктивный. Препродуктивный период включает следующие возрастные состояния: проростки, юные спорофиты и молодые спорофиты. В пределах репродуктивного периода выделяют два возрастных состояния: зрелого и стареющего спорофита. В пострепродуктивном периоде авторы выделяют лишь одно возрастное состояние старого спорофита.
Морфологическое описание спорофитов и гаметофитов папоротников проводили на натуральных объектах, при этом использовали терминологию, предложенную А.А. Федоровым с соавторами (1956, 1962). Измерение мелких объектов проводилось с помощью мерной лупы. Данные количественных показателей обрабатывались методами вариационной статистики (Зайцев, 1990). При этом определяли такие показатели, как: среднее значение признака (М=SX/N), ошибка среднего значения (m=s/![]() ), среднее квадратическое отклонение (s=
), среднее квадратическое отклонение (s=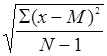 ), коэффициент вариации (V=s·100/M).
), коэффициент вариации (V=s·100/M).
Для оценки амплитуды изменчивости использовали унифицированную шкалу уровней изменчивости, разработанную С.А. Мамаевым (1969): V меньше 7% - очень низкий, V = 8-12% - низкий, V = 13-20% - средний, V = 21-30% - повышенный, V = 31-40% - высокий и V больше 40% - очень высокий уровень изменчивости.
Глава III.
Ботаническая характеристика объектов исследования
3.1. Общая характеристика отдела Папоротниковидные (Polypodiophyta). Папоротниковидные относятся к числу наиболее древних групп высших растений. По своей древности они уступают только Риниофитам и Плауновидным и имеют приблизительно один возраст с Хвощевидными. Риниофиты – это вымершая группа растений; Плауновидные и Хвощевидные играют в современном растительном покрове Земли очень скромную роль; и лишь Папоротниковидные в настоящее время продолжают процветать. Современные ботаники насчитывают около 300 родов и более 10 000 видов папоротников (Тахтаджян, 1978а; Рейвн и др., 1990).
Папоротники распространены очень широко, фактически по всему земному шару, и встречаются в самых различных местообитаниях, начиная с пустынь и заканчивая болотами, озерами, рисовыми полями и солоноватыми водами. Но наибольшее их разнообразие наблюдается в тропических лесах (Ярошенко, 1975; Леме, 1976; Второв, Дроздов, 1978). Еще во второй половине XIX века Альфред Уоллес указывал на большое количество и разнообразие жизненных форм папоротников в экваториальной зоне. Так, только на одной из вулканических вершин на острове Яве насчитывается около 300 видов папоротников (Уоллес, 1975). Обилием папоротников удивляет флора Новой Зеландии, многие виды которых являются здесь эндемиками. Видовой эндемизм папоротников Новозеландской флористической области составляет 40%, а у родов Диксония (Dicksonia) и Циатея (Cyathea) – примерно 75% (Тахтаджян, 1978б). Очень часто встречаются здесь древовидные папоротники, которые придают лесу особый облик. Это прежде всего Циатея серебристая (C. dealbata), которая считается национальным растением Новой Зеландии (Петров, 1981). Богата видами птеридофлора Новогвинейской флористической области. Из 1000 видов папоротников, встречающихся здесь, около 600 являются эндемиками (Второв, Дроздов, 1978).
Полный жизненный цикл папоротников складывается из двух фаз – фазы гаметофита и фазы спорофита. Когда говорят о папоротниках, то имеют в виду прежде всего спорофит. Как и у большинства высших растений (за исключением Моховидных), спорофит папоротников является доминирующей фазой в их жизненном цикле.
Спорофит папоротников имеет корни, за исключением небольшого числа видов, что является результатом редукции. Корни у папоротников только придаточные, т.е. главный корень не получает здесь дальнейшего развития и вскоре отмирает. Впервые стержневая корневая система появляется лишь у голосеменных растений (Комаров, 1936).
Стебель у большинства современных папоротников представлен подземным корневищем, иногда мощным, иногда тонким, шнуровидным. У тропических древовидных форм он надземный, прямостоячий, одет массой отмерших листьев. Анатомическое строение стебля может быть весьма сложным и разнообразным. Проводящая система представлена протостелой, сифоностелой или диктиостелой. В эволюционном отношении диктиостела наиболее продвинутый тип и поэтому она характерна для наиболее прогрессивных семейств (Тахтаджян, 1978а).
Листья папоротников, как и большинства других высших растений, возникли в результате уплощения и срастания дихотомически ветвящихся теломов Риниофитов (Серебряков, 1952; Мейер, 1958; Ярошенко, 1963; Тахтаджян, 1978а; Письяукова, 1980). Убедительным доказательством веточной природы листьев является характерный для них верхушечный и длительный (иногда почти неограниченный) рост, большие размеры и сложно рассеченная листовая пластинка, крупные и сложные листовые следы и наличие листовых прорывов (лакун) в стеле. По морфологическому и анатомическому строению листья папоротников весьма разнообразны. Их размеры колеблются от нескольких миллиметров до 30 м в длину. В большинстве случаев листья папоротников совмещают функции фотосинтеза и спороношения. У некоторых папоротников (Страусник – Struthiopteris, Оноклея – Onoclea) листья дифференцированы на стерильные (фотосинтезирующие) и фертильные (несущие спорангии). Чаще всего листья папоротников перистые, хотя имеются виды (Пиррозия длинночерешковая – Pyrrosia petiolasa, Щиточешуйник уссурийский – Pleopteris ussuriensis, Кривокучник сибирский – Camptosorus sibiricus, Листовик обыкновенный – Phyllitis scolopendrium), листья которых цельные. Пластинка перистого листа имеет стержень (рахис). Если лист однажды перистый, то сегменты листа называются перьями. Сегменты второго, третьего и других порядков называются перышками.