Исследование расщепления крахмала под действием a-амилазы слюны
Биологически активное вещество в дальнейшем было выделено из солода и подверглось детальному изучению. Было обнаружено, что его действие на крахмал происходит через три стадии: разжижение, декстринизацию и осахаривание. Это привело к признанию существования в солоде двух различных компонентов, которые в дальнейшем были получены в отдельности и названы a-амилазой (декстринирующий компонент) и b-амилазой (осахаривающий компонент).
a-Амилазы и b-амилазы широко распространены в высших растениях. Наиболее важным источником амилаз являются хлебные злаки, зерно которых в проросшем состоянии (в виде солода) находит широкое применение в промышленном гидролизе крахмала. Солод из ячменя, ржи, пшеницы, овса, проса в настоящее время используются для осахаривания крахмала в спиртовом производстве.
3.4. a-Амилазы
Роль a-амилаз при гидролизе крахмала исключительно велика. Из трёх основных функций при действии на клейстеризованный крахмал (разжижение, декстринизация, осахаривание) разжижение и декстринизация зависят от a-амилаз. Они атакуют не только клейстеризованный, но и нативный крахмал, разрушая крахмальные зёрна.
a-Амилазы различного происхождения имеют много общих свойств: хорошо растворяются в воде или в сильно разбавленных растворах солей. Более концентрированные растворы солей (например 20-30% - ные растворы сульфата аммония) вызывают осаждение этих ферментов. a-Амилазы легко растворяются в разбавленных растворах этилового спирта, но осаждаются при его концентрации в среде свыше 60%. Белок a-амилаз обладает слабокислыми свойствами; изоэлектрическая точка ферментов колеблется в пределах рН 4,2 – 5,7. Молекулярная масса солодовой a-амилазы 60000, a - амилаз микроскопических грибов – 45000-50000. Многие из известных a-амилаз получены либо в высокоочищенном, либо в кристаллическом виде.
Ионы кальция оказывают стабилизирующее действие на a-амилазы. Это впервые было обнаружено Воллерштейном, затем потверждено Накамурой. В настоящее время это явление отмечено почти для всех амилаз. Однако теоретически этот вопрос применительно к промышленному гидролизу крахмала до сих пор не разработан.
3.5. Гидролиз крахмала a-амилазами
a-Амилазы действуют на a-1,4 –гликозидные связи, расщепляют амилозу внутри её цепи, в результате чего с большой скоростью образуются низкомолекулярные продукты гидролиза – нормальные a-декстрины. Их дальнейший гидролиз даёт мальтозу, мальтотриозу и глюкозу. Было найдено, что расщепление a-1,4-глюкозидных связей в амилозе носит случайный характер иподчиняется закону статистического распределения продуктов реакции. Расщепление более мелких фракций на последнем этапе амилоза носит уже не случайный характер – действие фермента направлено лишь на определённые a-1,4-гликозидные связи. В конечном счёте a-амилазы превращают амилозу в мальтозу и глюкозу, хотя и отмечены некоторые несущественные различия в динамике гидролиза этими ферментами указанного субстрата.
Бендецкий, Яровенко по изменению вязкости и восстанавливающей способности гидролизатов крахмала оценивали действие (множественность атаки) a-амилазы Bac. Subtilis на растворимый крахмал. Авторы наблюдали существенное различие вязкость –восстанавливающая способность при кислотном и ферментативном гидролизе крахмала. Это дало основание сделать заключение, что при кислотном гидролизе деградация крахмала происходит беспорядочно, а при действии a-амилазы осуществляется множественная атака на субстрат, приводящая к образованию олигомеров на первой стадии деградации.
Разрыв цепей амилопектина осуществляется между a-1,6 - глюкозидными связями. Отщепление декстринов, содержащих 15 и более гликозидных остатков, идёт с большей скоростью, в то время как конечное осахаривание существенно замедляется. Продукты гидролиза, содержащие a-1,4 - глюкозидные связи, являются нормальными a-декстринами и состоят из 6-13 глюкозидных остатков. Остаточные декстрины, содержащие большое количество a-1,6-связей, обазначаются как аномальные конечные декстрины. Установлено, что a-1,6-связи не только не расщепляются a-амилазой, но и являются стерическим препятствием для гидролиза a-1,4-связей, находящихся в непосредственной близости к a-1,6-связям. Наименьшим предельным декстрином в конце гидролиза амилопектина амилазой слюны был найден тетрасахарид, солодовой амилазы – паноза, т. е устойчивым к расщеплению в первом случае были две a-1,4-глюкозидные связи, во втором – лишь одна.
На рис. Представлена схема действия a-амилаз на амилозу и амилопектин по Бернфельду.
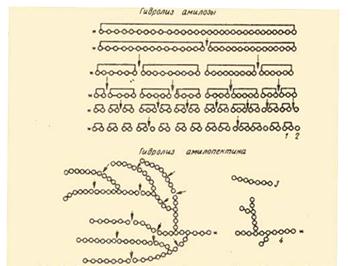
![]() Рис.2 Действие a-амилазы на амилозу и амилопектин по Бернфельду.1 – мальтоза; 2 – глюкоза; 3 – нормальный a-декстрин; 4 – конечный a-декстрин; *-редуцирующий конец амилозы или амилопектина; - действие a-амилазы.
Рис.2 Действие a-амилазы на амилозу и амилопектин по Бернфельду.1 – мальтоза; 2 – глюкоза; 3 – нормальный a-декстрин; 4 – конечный a-декстрин; *-редуцирующий конец амилозы или амилопектина; - действие a-амилазы.
4. Ход работы
Оборудование: электрическая плитка, химические стаканы, колбы, термостойкая колба, термометр, водяная баня, пипетка, мензурка.
Реактивы: крахмал, раствор йода.
Приготовление раствора крахмала:
В термостойкую колбу добавляем 200 мл воды. В неё же добавляем 2 г крахмала. Доводим смесь до кипения и кипятим несколько минут. Таким образом получаем 1% раствор крахмала.
Приготовление раствора слюны:
50 мл воды полощем во рту в течении 3-4 минут.
Проведение эксперимента:
В три пробирки добавляем по 25 мл раствора крахмала и каплю йода. Раствор крахмала окрашивается в интенсивно синюю окраску. Нагреваем электрическую плитку и измеряем температуру водяной бани. В течении 8-10 минут даём время, чтобы температура водяной бани и температура раствора в пробирках уравнялась. Снова измеряем температуру водяной бани. Добавляем в раствор крахмала 5 мл раствора слюны и включаем секундомер. Отмечаем время исчезновения синей окраски. Затем в следующие три пробирки добавляем раствор крахмала и каплю раствора йода, охлаждаем водяную баню и повторяем процедуру при более низкой температуре.
Результаты эксперимента приведены в таблице:
|
toC |
22 |
30 |
34 |
37 |
40 |
45 |
58 | |
|
t1 |
11 мин 17 с |
7 мин 5с |
3 мин 56 с |
3 мин 16 с |
4 мин 6 с |
5 мин 13 с |
6 мин 57 с | |
|
t2 |
11мин 22 с |
7 мин 13с |
4 мин 6 с |
3 мин 23 с |
4 мин 16 с |
5 мин 5 с |
6 мин 55 с | |
|
t3 |
11 мин 23 с |
7 мин 9с |
4 мин |
3 мин 21 с |
4 мин 8 с |
5 мин 11 с |
7 мин 3 с | |
|
tср. |
11 мин 21с |
7 мин 9 с |
4 мин 1 с |
3 мин 20 с |
4 мин 10 с |
5 мин 10 с |
6 мин 58 с | |
|
toC |
22 |
30 |
34 |
37 |
40 |
45 |
58 | |
|
1/t мин-1 |
0.088 |
0.14 |
0.25 |
0.3 |
0.24 |
0. 195 |
0.143 | |