Методы разделения иммуноглобулинов
ВВЕДЕНИЕ
Иммуноглобулины - основные защитные белки организма, так как они обладают свойствами различных антител. Они содержатся в крови, молозиве и молоке, слюне и других жидкостях. С их количеством и активностью связаны жизнестойкость, физиологическое состояние и продуктивность животных.
В работе описаны свойства иммуноглобулинов, как белков, а также способы разделения иммуноглобулинов сельскохозяйственных животных.
Представленные данные по характеристике, выделению иммуноглобулинов, могут дать современные представления о защитных белках животных.
ИММУНОГЛОБУЛИНЫ - СПЕЦИФИЧЕСКИЕ ФАКТОРЫ ГУМОРАЛЬНОГО ИММУНИТЕТА
Важнейшие факторы специфического иммунитета - иммуноглобулины, осуществляют гуморальную защиту организма.
Дж.Хереманс (1959), изучая активность иммунной сыворотки человека, обнаружил, что антитела содержатся, по меньшей мере, в трех фракциях сыворотки: в g- и в двух фракциях b- глобулинов, называемых b2А и b2М - глобулинами. Эти фракции, активные в иммунологическом отношении, Хереманс предложил назвать иммуноглобулинами (Ig). Предложение Дж.Хереманса легло в основу международной номенклатуры, принятой в 1664 г. специальной комиссией ВОЗ. По этой номенклатуре g-глобулин называется IgG, B2A-глобулин - IgA и В2М - IgM. Иммуноглобулин состоит из двух легких (L) и двух тяжелых (Н) полипептидных целей. Причем каждый класс иммуноглобулинов характеризуется особым типом тяжелых цепей g, m, a, d, e.
Легкие цепи бывают двух типов - k или l, каждвй из которых отличается С- концевой последовательностью аминокислот. Как тяжелые, так и легкие цепи состоят ив двух различных участков (областей) - вариабельной (V) и постоянной (С). С-концевая половина полипептидной цепи имеет постоянную аминокислотную последовательность, а ее N - концевая часть - вариабельную. Каждый постоянный (Сl) и вариабельный (Vl) участок легкой цепи включает 107-110 аминокислотных остатков.
Тяжелые цепи построены их четырех участков - VH, C1H, C2H, C3H. Вариабельные участки их состоят приблизительно из 110-114 аминокислотных остатков, постоянные - 330.
В вариабельной части полипептидных цепей находятся определенные, так называемые "гипервариабельные участки", с наибольшим числом аминокислотных замен. В легких цепях они расположены между 24-34; 52-55; 89-97 аминокислотными остатками. Гипервариабельные участки тяжелых цепей занимают аналогичные положения, но точная локализация их пока не установлена (Кульберг, 1975; Поляк 1981).
Наличие вариабельных участков дает молекулам антител возможность приспосабливаться к разнообразным антигенным детерминантам. Строение постоянных областей тяжелых цепей определяет эффекторные функции молекул к поверхностям макрофагов. В-лимфоцитов, тучных клеток, а также проникновение через плацентарную мембрану (Егоров , 1972; Leslie, Cohen, 1973).
Гибкость молекул иммуноглобулинов, обеспечивающая приспособляемость к различным конфигурациям молекул антигена, обусловливается также наличием особого "шарнирного участка" в середине тяжелых цепей, содержащего много остатков аминокислоты пролина и препятствующего образованию вторичной структуры.
В настоящее время на основании изучения первичной структуры полипептидных цепей выдвинута так называемая "доменная" гипотеза строения иммуноглобулинов, согласно которой молекулу иммуноглобулинов можно разбить на участки с относительно независимыми конфигурациями в виде глобул. Каждый домен состоит приблизительно из 100-110 аминокислотных остатков и имеет одну дксудьфидную связь, которая связывает участки цепей, образуя петлю из 60 аминокислотных остатков (Еdelman, 1970; Незлин, Рохлин, 1970; Роljak, 1972).
Молекула иммуноглобулина G состоит из 12 доменов: по 4 в тяжелых и по 2 в легких полипептидных цепях (рис.1). Используя молекулярные модели и ряд общих термодинамических принципов укладки третичной структуры белков, установлено, что конформации всех 12 доменов иммуноглобулина G характеризуются большим количеством b- складок и наличием одно-двух витков a- спиральной структуры (Завьялов, Троицкий, 1973).
На основании изучения воздействия протеолитических ферментов сделан вывод, что молекулы иммуноглобулинов состоят из в трех фрагментов двух типов, связанных гибкими участками полипептидных цепей: два из них названы Fab- и один Fc- фрагментами (Рогtег, 1959, 1960; Utsumi Karush, 1955, 1967).
В состав FAB- фрагментов входят целая легкая цепь и N - концевая половина тяжелой цепи. Структурная особенность FAB- фрагмента наличие двух глобулярных субъединиц - V и С. V построена из двух доменов VH и V1L, а С- из CL и C1H. Fc-фрагмент содержит по два домена - C2H и Сн3 (Роljak, 1972).
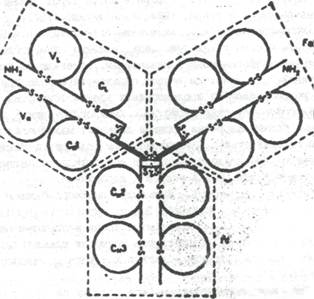 |
Рис. 1. Схема строения молекулы IgG1 человека (из книги Иммуноглобулины".- М.: Мир., 1981).
L- цепи разделены на две гомологичные области VI. (вариабельная) и СL (константная). Утолщенные линии Н-цепей соответствуют шарнирному участку. Показаны 4 гомологичные области Н-цепей (VF, С1Н, С2Н, С3Н), внутри - и межцепочные дисульфидные связи, N- концевые участки обоих цепей и основные фрагменты (FAB, FAB' и Fc).
В состав молекул некоторых классов иммуноглобулинов входят дополнительно полипептидные цепи. В молекуле сывороточного и секреторного иммуноглобулина А она названа J- цепочкой. (НАLPERN, KOSHLAND, 1970). Выяснено, что аналогичные полипептиды содержатся также и в молекулах иммуноглобулина М (Меstresky et al., 1971). J-цепочки иммуноглобулинов М и А имеют одинаковые молекулярную массу, электрофоретическую подвижность, антигенную специфичность и аминокислотный состав (Меtresky et al, 1971; Моrrison, Коshland, 1972). В полимерных молекулах иммуноглобулинов различных видов животных также содержится полипептид, аналогичный полипептиду J- цепочки иммуноглобулинов человека (Kobajashi et al, 1973).
Молекулярная масса J- цепи колеблется от 14 000- 27000. Такое расхождение объясняется, по-видимому, склонностью J цепей образовать димеры (Коshland, 1975). Содержание углеводов в них около 7,5 моль %. Определен аминокислотный состав J- цепей выделенных из IgA и IgМ человека, а также животных. Изолированные цепи отличаются высоким содержанием аспарагиновой, глутаминовой кислот и цистеина и почти полным отсутствием трипто-фана (Morrison, Коshland, 1972; Zikand 1972; Kehoe et al, 1972; Mestesky et al 1975).
Предполагается, что J цепи необходимы для присоединения секреторного компонента к IgA и облегчают димеризацию IgA b и полимеризацию IgМ. По мнению Л.В.Часовниковой и др. (1983), большая информационная стабильность молекул IgA, по сравнению с IgM обусловлена наличием J- цепи, объединяющей молекулу IgA в димерную форму.
В состав секреторного IgА входит еще один полипептид, названный секреторным компонентом (Brandtzaeg, 1968, 1974; Fomasi et al, 1968). Секреторный компонент обнаружен у всех видов сельскохозяйственных животных, коров (Mashetal, 19б9; Butler et al, 1972); свиней (Bourle, 1969); кроликов (Halperl, Koshland, 1970); овец и коз (Pahud Mach, 1970); лошадей (Pahud Mach, 1972). Он существует как в связанном с секреторным IgA - (сСК), так и в свободном виде - (нСК). Молекулярная масса связанного и свободного секторного компонентов у человека колеблется от 50 000 до 90 000, у овец, коз и лошадей молекулярная масса нСК 80 000- 85 000 (Pahud Mach, 1970, 1972). Содержание углеводов - 15,9 - 22,8%. Определен также аминокислотный состав свободного секреторного компонента у человека, крупного рогатого скота и собаки (O’Daily, Cerba, 1971; Fomasi, Jrey, 1972; Mestesky, 1972; Fhopson et al, 1975).