Бактериальная система секреции белков первого типа
Система секреции α-гемолизина Escherichia сoli представляет собой прототип системы секреции первого типа, и на сегодняшний день хорошо изучена. Она состоит из трех компонентов: TolC, HlyD, HlyB. Белок TolC является аналогом OMP для экспорта α-гемолизина, и представляет собой тримерный комплекс, расположенный во внешней мембране. Предполагается, что он состоит из пориноподобного β-складчатого мембранного домена с гидрофильной карбокси-концевой областью, расположенной в периплазматическом пространстве. Однако, недавний анализ последовательности указывает на то, что TolC и другие OMP не являются поринами. OMP функционирует как канал секреции через внешнюю мембрану, что было доказано порообразующим действием олигомеров TolC в экспериментальных липидных бислоях (D. Thanassi еt al., 2000). Периплазматический MFP (HlyD) также является тримерным и взаимодействует и с OMP, и с ABC-транспортером (HlyB). HlyD содержит короткий гидрофильный амино-концевой домен, заякоренный во внутренней мембране, включающий около 150 аминокислотных остатков; крупный гидрофобный домен, расположенный в периплазме, включающий 275 аминокислотных остатков, и карбокси-концевой домен, имеющий β-складчатую структуру, способный связываться с внешней мембраной, содержащий 275 аминокислотных остатков (M. J. Fath еt al., 1993). Предполагается, что MFP облегчает секрецию субстрата без промежуточного периплазматического звена, формируя закрытый канал, соединяющий внутреннюю и внешнюю мембраны, и осуществляя прямой контакт между ABC-транспортером и OMP. Что касается HlyB, то его точное строение пока не установлено, предполагается, что он состоит из восьми доменов. Два из них в амино-концевой области и шесть в центральной гидрофобной области. Результаты экспериментального изучения этого аппарата привели к возникновению двух моделей секреции первого типа (D. Thanassi еt al., 2000).
Эксперименты по секреции α-гемолизина E. coli показывают, что ABC-транспортер и MFP ассоциируются еще до связывания с субстратом. Прикрепление субстрата к этому комплексу вызывает контакт MFP с OMP. Это соединение является обратимым, и разрушается сразу после экспорта субстрата. Энергия гидролиза АТФ посредством ABC-транспортера расходуется только на транслокацию субстрата и не требуется для связывания субстрата или для сборки комплекса (D. Thanassi еt al., 2000).
Эксперименты по секреции гемопротеина Serratia мarcescens и металлопротеазы Erwinia chrysanthemi указывают на немного иной порядок событий. По этой модели, ABC-транспортер и MFP не связываются перед закреплением субстрата. Субстрат в первую очередь связывается с ABC-транспортером, затем образовавшийся комплекс ассоциируется с MFP, и только потом происходит связывание с OMP, после чего происходит секреция субстрата. Для определения правильной модели, или для уточнения возможных индивидуальных отличий в функционировании аппарата секреции первого типа необходимы дальнейшие исследования (D. Thanassi еt al., 2000).
Было установлено, что ОМР системы секреции α-гемолизина (TolC), используется также в системе секреции колицина V и в некоторых других системах, например при сегрегации хромосом, а также он может формировать канал во внешней мембране, специфический для медикаментов. ОМР системы секреции гемопротеина S. marcescens, называемый HasF, является в высокой мере идентичным с TolC E. сoli. Для воссоздания секреции HasА у E. сoli необходимо наличие в качестве ОМР либо TolC, либо HasF, либо PrtF. Такие гибридные секреторные системы функционируют как для секреции HasA, так и для секреции протеазы. Это является типичным примером комплементации ОМР (R. Binet et al., 1997). В частности, степень гомологии между компонентами системы секреции липазы S. marcescens, белками lipB, lipC, lipD и компонентами транспортера металлопротеазы Er. chrysanthemi PrtD, PrtE, PrtF составляет 45-55%. А гомология между LipB и LipC, и HasD, и HasE у S. marcescens составляет 45-53%. Эти показатели считаются довольно высокими (H. Akatsuka et al., 1998). Однако было выявлено, что не все комбинации между компонентами гибридных секреторных систем являются активными. Так, HasE формирует активные экспортеры и с PrtF, и с TolC, тогда как PrtE может формировать активный экспортер только с PrtF, но не с TolC. Исследования этих мультибелковых комплексов in vitro подтвердили существование некоторых функциональных различий между HasE и PrtE. Полученные результаты могут быть полезными при определении сайтов, ответственных за связывание MFP и OMP (H. Akatsuka et aj., 1998).
С другой стороны, исследования in vivo и in vitro показывают, что HasD и PrtD могут образовывать активные секреторные системы с PrtE и HasE в любых комбинациях (H. Akatsuka et al., 1998).
Также были проведены исследования по изучению секреции липазы LipA S. marcescens посредством системLipB-LipC-LipD и HasD-HasE-HasF. В результате опытов было выяснено, что HasD-HasE-HasF-транспортер осуществляет секрецию LipA так же эффективно, как и LipB-LipC-LipD. LipB-HasE-HasF-система могла производить секрецию LipA, но не была способна секретировать HasA, система HasD-Lip-CLipD не была способна к секреции обоих субстратов (H. Akatsuka et al., 1998).
В случае экспериментов с системами секреции липазы LipA S. marcescens и металлопротеазы PrtC E. chrysanthemi были получены сходные результаты, приведенные в таблице:
Таблица 1
Эффективность гибридных систем секреции (по H. Akatsuka et al., 1998).
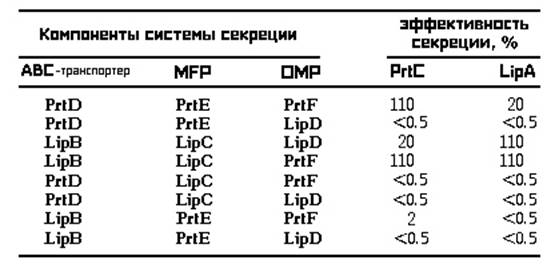
Не все комбинации компонентов привели к формированию эффективных систем секреции. Полученные результаты позволили сделать некоторые конкретные выводы. В частности, что PrtD-PrtE-LipD-система не способна экспортировать ни LipA, ни PrtC, в то время как, LipB-LipC-PrtF-система оказалась настолько же функциональной для LipA секреции в E. coli как и в S. marcescens. PrtE может взаимодействовать только с PrtF, тогда как HasE и LipC показывают более широкие возможности связывания с различными белками. Было также установлено, что PrtD не может ассоциироваться с LipC, а LipB-PrtE-PrtF-система является очень неэффективной в отношении экспорта LipA и PrtС (H. Akatsuka et al., 1998).
В ходе исследований было установлено влияние шаперона SecB на процесс секреции HasA у S. marcescens. Точное его значение на данный момент не установлено, но было показано, что инактивация этого шаперона приводит к блокированию секреции HasA (P. Delepelaire et al., 1998).
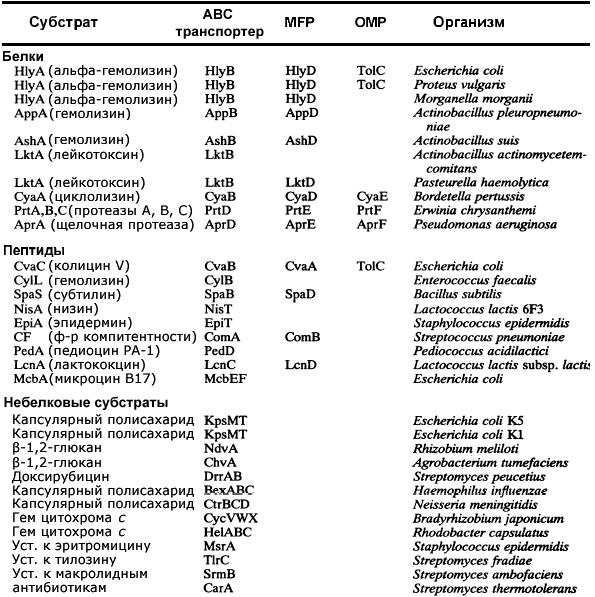
К настоящему времени установлено строение систем секреции первого типа у многих микроорганизмов, некоторые из них приведены в таблице 2. Однако остается довольно большое количество секреторных систем неполного состава, для которых остаются невыясненными либо некоторые компоненты, либо субстраты (M. J. Fath еt al., 1993).
Таблица2
Некоторые системы секреции первого типа (по M. J. Fath еt al., 1993).
ABC-транспортеры