Биогенез мембран
Оглавление
1 Введение
2 Образование и встраивание мембранных белков
2.1 Транспорт белков
2.2 Сигнальные последовательности белков
2.3 Стоп- сигналы переноса
2.4 Сигнальные пептидазы
2.5 Белки необходимые для распознавания сигналов переноса
3 Липиды
3.1 Синтез у прокариот
3.2 Синтез у эукариот
3.3 Транспорт липидов
3.3.1 Транспорт у прокариот
3.3.2 Транспорт липидов у эукариот
3.4 Изменение в липидном составе под действием окружающей среды
4 Заключение
5 Список литературы
1 Введение
Ранние этапы становления жизни являются неизученными проблемами биологии. На сегодня ни одна из многочисленных теорий не в состоянии дать устраивающего всех варианта происхождения жизни.
Идеальные условия для образования и сколь-нибудь длительного существования нужных для биогенеза молекул могли быть созданы только при наличии комфортной среды, которая отличалась бы от агрессивного окружения. По всей вероятности такие условия были созданы при появлении липидов, которые при взаимодействии с водой образовали примитивные липосомальные микросферы. По своей пространственной организации замкнутая форма липидной мембраны соответствует наименьшему значению энергии Гиббса, то есть термодинамически выгодна по сравнению с другими возможными формами расположения молекул. Кроме того, конформационная специфика бислойной фосфолипидной оболочки соответствует жидкокристаллическому состоянию, что предусматривает автономность по отношению к окружающей среде и одновременно селективную и регулируемую связь с этим внешним окружением.
Этот уникальный вариант не мог не закрепиться в ходе последующей биологической эволюции и не создать предпосылок для формирования гомеостаза, как одного из основополагающих принципов жизни.
Биогенез мембраны начинается с процессов синтеза составляющих ее компонентов – белков, липидов, углеводов. Затем эти компоненты должны быть доставлены к месту назначения и там образовать нужные структуры.
2 Образование белков
Две главные проблемы касающиеся сборки мембранных белков.
1. Все закодированные в ядре белки синтезируются общим пулом рибосом. В связи с этим возникает вопрос: как отдельные мембранные белки доставляются к месту назначения? Чем отличаются белки плазматической мембраны от белков внутренней митохондриальной мембраны или от белков мембран эндоплазматического ретикулума? Эту сложную проблему сортировки можно решить только при наличии определенных сигнальных последовательностей в каждом полипептиде, а также соответствующего аппарата узнавания.
2. Каков истинный механизм встраивания мембранных белков в мембрану и как при этом достигается правильная их ориентация относительно мембранного бислоя? Требуют ли механизмы встраивания и ориентации также наличия определенных сигнальных элементов и систем узнавания и если да, то каковы они? Какие свойства обеспечивают при встраивании мембранных белков формирование правильной третичной, а также червертичной структуры в случае мультисубъедииичных ансамблей?
В поиске ответов достигнуты большие успехи. Существует идентифицируемая часть полипептидной последовательности, которая служит участком узнавания, или «сигналом», направляющим отдельный полипептид к мембране, в которую он встраивается. Эти сигнальные участки часто расположены на N-конце новосинтезированного полипептида и отщепляются специфическими сигнальными пептидазами после встраивания его в нужную мембрану. Для обозначения N-концевого сигнала различными авторами использовались следующие термины: сигнальный пептид, сигнальная последовательность, транзитный пептид, лидирующий пептид, пре-последовательность.
Процессы трансляции и встраивания белков в мембрану можно разделить в эксперименте. Для сборки мембранных белков в большинстве случаев необходима энергия, отличающаяся по величине от той, которая требуется для их трансляции на рибосоме. Связавшийся с мембраной-мишенью полипептид должен, кроме того, находиться в конформации, в которой может осуществляться его перенос через мембрану или встраивание в нее. Во многих случаях перенос белков через мембраны происходит от N-конца к С-концу, при этом необходимо, чтобы белок был по крайней мере частично развернут или слабо свернут.
Доставка каждого белка к месту назначения обеспечивается иерархией сигналов, закодированных в каждом полипептиде. Например, большинство белков, предназначенных для эндоплазматического ретикулума или митохондрий, синтезируется в виде предшественников большей молекулярной массы (пре-белков); на N-конце у них имеется дополнительная последовательность, которая отщепляется особыми протеолитическими ферментами, имеющимися в этих орган ел л ах. Такие первичные сигналы весьма разнообразны и необходимы для того, чтобы полипептиды были узианы при транслокации специфическими рецепторами в этих органеллах. Связывание с митохондриями происходит сразу после завершения трансляции. Однако для большинства белков, направляемых в
эндоплазматический ретикулум в клетках млекопитающих, наблюдается иная картина. Как видно из рис.1 после связывания белков с соответствующей органеллой должна произойти дальнейшая сортировка. Для этого нужна дополнительная информация, которая также должна быть закодирована в каждой полипептидной последовательности и может рассматриваться как вторичные сигналы. В нескольких случаях их удалось идентифицировать как сигнальные последовательности, физически отделенные от первичных, хотя, возможно, так бывает не всегда.
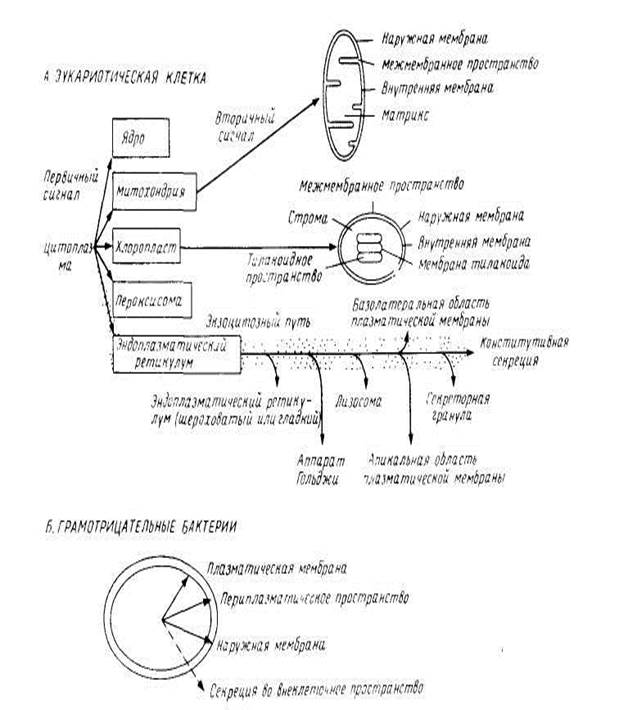
рис.1 Сортировка мембранных белков
биогенез мембрана белок липид
Особый интерес представляет процесс сборки мембранных белков, который целесообразно рассмотреть в связи с их сортировкой. На рис. 2 схематически показаны три общих механизма проникновения пептидного предшественника в мембрану. Механизмы А и Б являются вариантами схемы линейного вытеснения, согласно которой сигнальная последовательность направляет полипептид к переносящему устройству, которое включает в себя заполненный водой канал. Сигнальная последовательность может проходить прямо сквозь канал (механизм А) или оставаться связанной с мембраной, образуя, как показано на рис.2, петлю (механизм Б). В отсутствие какого-либо сигнала остановки процесса переноса полипептид будет транспортироваться через мембрану целиком. Однако, если внутри полипептида имеется второй сигнальный пептид, называемый стоп-сигналом переноса, то процесс останавливается и стоп-сигнал переноса становится трансмембранным сегментом зрелого мембранного белка. Фиксируя белок в мембране, стоп-сигнал переноса действует как сигнал сортировки. Если в белке имеются и другие сигналы начала и конца переноса, то будут образовываться следующие трансмембранные сегменты. Схема В на рис.2 иллюстрирует возможную роль самопроизвольного включения в мембрану гидрофобных элементов полипептидного предшественника. Этот механизм может реализовываться только тогда, когда включение в мембрану происходит после трансляции полипептида. Примером, подтверждающим существование этого механизма, является пробелок оболочки фага М13. Модель самопроизвольного включения может использоваться для объяснения механизма встраивания поперек мембраны амфифильных а-спиралей или β-структур. Этот процесс может также, конечно, быть белокзависимым.