Биологическая сущность микоризы
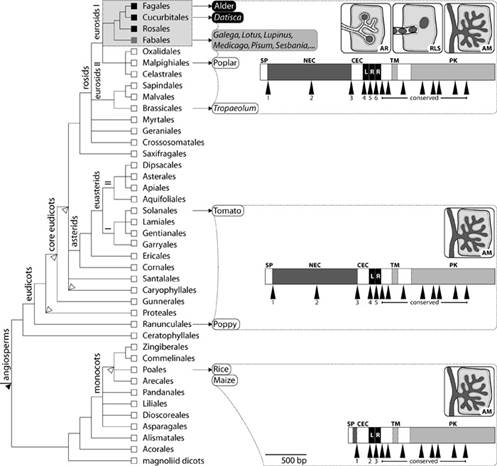
Рис. 2.1. Структура гена SYMRK и типы внутриклеточного корневого симбиоза у цветковых растений. Слева— эволюционное древо цветковых. Серым прямоугольником обозначены четыре порядка, у представителей которых встречаются клубеньковые симбиозы (АР или СБР). В овальные рамки заключены родовые названия растений, у которых анализировалась структура гена. цветом выделены растения, образующие АМ и АР, серым— АМ и СБР, белым— только АМ. Схематичные рисунки изображают три типа внутриклеточного симбиоза: AR=АР, RLS=СБР, AM=АМ. Три длинных горизонтальных прямоугольника отображают структуру гена SYMRK. Буквами обозначены домены: NEC, LRR— предполагаемые рецепторные (внеклеточные) домены; TM— трансмембранный домен; PK— протеин-киназный (внутриклеточный) домен. (Рис. из обсуждаемой статьи вPLoS Biology )
Обнаружив эту закономерность, авторы, естественно, предположили, что приобретение "длинного" варианта SYMRK и было тем самым ключевым событием, которое создало необходимые предпосылки для развития клубеньковых симбиозов с бактериями— причем "генетическая программа" клубенькового симбиоза представляет собой модификацию "генетической программы" арбускулярной микоризы. Чтобы подтвердить или опровергнуть это предположение, ученые провели серию генно-инженерных экспериментов.

Рис. 2.2. Растение Datisca glomerata (Katharina Markmann, 2008)
Первый эксперимент показал, что ген SYMRK необходим не только для АМ и СБР (что было известно и ранее), но и для АР. Ученые отключили ген SYMRK у растения Datisca glomerata, корни которого в норме образуют АМ и АР (рис. 2.2). результате растение практически полностью утратило способность к формированию обоих симбиозов— и с грибом, и с актинобактерией Frankia. Тем самым впервые удалось показать, что ген SYMRK необходим для всех трех типов внутриклеточного симбиоза: АМ, СБР и АР. Стало ясно, что АР имеет, скорее всего, примерно ту же генетическую "основу", что и СБР ( ранее о генетики АР не было известно практически ничего).
Второй эксперимент показал, что ген SYMRK не служит для распознавания конкретных бактерий-симбионтов. Бобовое растение лядвенец японский (Lotus japonicus) образует СБР с бактерией Mesorhizobium loti, алюцерна (Medicago truncatula)— с бактерией Sinorhizobium melioti. Мутантной люцерне, имеющей "испорченный" ген SYMRK и не способной формировать ни АМ, ни СБР, пересадили "здоровый" ген SYMRK от лядвенца. Эта операция полностью восстановила способность люцерны образовывать оба типа симбиоза. При этом трансгенная люцерна стала образовывать СБР со "своей" исконной бактерией Sinorhizobium, а вовсе не с Mesorhizobium. Таким образом, SYMRK не отвечает за распознавание и выбор бактериального симбионта, а только за общую способность формировать внутриклеточный симбиоз с бактериями. Распознавание осуществляется другими белками, какими именно— пока не установлено.
Третий эксперимент показал, что для обеспечения нормального клубенькового симбиоза, равно как и микоризы(АМ), вполне подходит любой "длинный" вариант гена SYMRK, взятый хоть у бобового растения, образующего СБР, хоть у растения с АР, хоть у вида, вовсе не образующего клубеньков. Для эксперимента были использованы мутантные растения Lotus japonicus, у которых ген SYMRK кодирует нефункциональный белок с испорченным киназным доменом. Эти растения не могут образовывать ни СБР, ни АМ. Им пересаживали гены SYMRK от разных растений, имеющих длинный вариант этого гена: от других бобовых, образующих СБР (люцерны), от актиноризных растений (Datisca glomerata) и, наконец, от бесклубеньковых родственников (настурции Tropaeolum majus). Все эти операции привели к полному восстановлению у мутантного лядвенца АМ и СБР (разумеется, со"своей" бактерией Mesorhizobium).
Четвертый эксперимент показал, что укороченные варианты гена SYMRK достаточны для АМ, но не для клубеньковых симбиозов. Как и в третьем эксперименте, использовали мутантную форму лядвенца японского, не образующую ни АМ, ни СБР. Растениям пересаживали "средний" вариант гена, взятый у помидора, и "короткий" вариант, позаимствованный у риса. В обоих случаях у мутантного лядвенца восстановилась способность к формированию АМ, но не СБР. На основе этих и ряда других экспериментов и наблюдений ученые заключили, что белок SYMRK, по-видимому, необходим для формирования особых внутриклеточных структур— своеобразных "с имбионтоприемников" или "пре-инфекционных нитей" (pre-infection threads), которые впоследствии заселяются симбиотическими бактериями (и тогда их уже называют "инфекционными нитями"). Похожие "симбионтоприемники" образуются в клетках корней и перед принятием грибных симбионтов при формировании АМ (эти структуры называются pre-penetration apparatus). Сходство в строении и механизмах формирования этих "симбионтоприемников", по-видимому, отражает единство генетической программы, отвечающей за формирование всех трех типов внутриклеточного симбиоза: АМ, СБР и АР. Мутации в некоторых "общих генах симбиоза" приводят к нарушению формирования "симбионтоприемников" (SYMRK, конечно, не единственный ген, необходимый для их формирования). Следует подчеркнуть, что "длинная" версия белка SYMRK, очевидно, является необходимым, но недостаточным условием формирования клубеньковых симбиозов. Это видно из того, что такие симбиозы могут формировать не все, а только некоторые обладатели "длинной" версии. По-видимому, растения, образующие клубеньки, должны обладать еще какими-то генетическими особенностями, которые пока не удалось обнаружить.
В целом, однако, полученные результаты убедительно подтверждают гипотезу, согласно которой способность к формированию клубеньковых симбиозов (АР и СБР) развилась на основе древней генетической программы АМ. Ключевое эволюционное событие заключалось в том, что клетки корней приобрели способность реагировать формированием "симбионтоприемников" не только на присутствие симбиотических грибов, но и на близость азотфиксирующих бактерий. Очень похоже, что в основе этого события лежало изменение структуры белка SYMRK, а именно добавление двух новых рецепторных доменов к его внеклеточной части. Оба эти домена могли быть заимствованы у генов других белков, имеющихся в геноме высших растений. Таким образом, возникновение клубеньковых симбиозов— яркий пример формирования новой функции путем модификации генного комплекса, ранее служившего для иных целей. Арбускулы— видоизменения мицелия у грибов - микоризообразователей, аналогичные гаусториям. Являются многократно дихотомически разветвлёнными гифами сложной формы, проникающими в паренхимальные клетки корня. Окружены внутри клетки клеточной плазмалеммой. В арбускулах происходит наиболее интенсивный обмен метаболитами между компонентами микоризы, хотя они и существуют лишь несколько дней (в последствии растворяются). Исследователями считается, что арбускулы образуются под влиянием защитной реакции клеток растения (Katharina Markmann и др., 2008)