Биполярные, горизонтальные и амакриновые клетки
Сигналы и рецептивные поля биполярных клеток зависят от двух механизмов. Во--первых, постоянное высвобождение медиатора в темноте способствует поддержанию ряда биполярных клеток в состоянии постоянной деполяризации, а других - в состоянии гиперполяризации, в зависимости от того, какие - возбуждающие или тормозные - рецепторы они имеют. Во-вторых, свет приводит к гиперполяризации фоторецепторов, тем самым уменьшая выброс глутамата. Уменьшение постоянного освобождения глутамата фоторецепторами при освещении приводит, соответственно, к уменьшению активности биполярных клеток, имеющих возбуждающие рецепторы глутамата, т.е. вызывают их гиперполяризацию. Такие клетки называются Н-биполяры (hyperpolarizing).
Наоборот, уменьшение тонического высвобождения глутамата фоторецепторами при освещении последних приводит к деполяризации тех биполярных клеток, которые экспрессируют тормозные рецепторы глутамата - это D-биполяры (depolarizing). D-биполярные клетки являются одним из редких типов клеток, у которых были обнаружены тормозные эффекты, опосредуемые через рецепторы глутамата. Канеко с коллегами показали, что данный тормозный эффект обусловлен метаботропными рецепторами глутамата, расположенными на мембранах биполярных клеток, которые действуют через G-белок и вторичные посредники, приводя к уменьшению ионной проводимости. Таким образом, в темноте в результате активации этих рецепторов ионные каналы, управляемые циклическими нуклеотидами на D-биполярных клетках, закрываются (в отличие от каналов фоторецепторов). После освещения высвобождение глутамата уменьшается, открываются катионные каналы, и биполярные клетки деполяризуются.
Структура рецептивных полей биполярных клеток
Рецептивное поле гиперполяризующейся Η-биполярной клетки показано на рис.4. Небольшой пучок света, направленный на центральную область поля, вызывает длительную гиперполяризацию. Освещение в форме кольца с темной зоной в центре приводит к деполяризации. Таким образом, рецептивная область подобной клетки состоит из различных по светочувствительности центральной зоны и окружающей ее периферии. Н-биполярная клетка, показанная на рис.4, может быть описана как клетка, имеющая рецептивное поле с "off"-центром, поскольку она деполяризуется, когда поток света исчезает.
D-биполярная клетка имеет рецептивное поле сходной концентрической формы, но освещение центра ее поля приводит к деполяризации, а освещение в форме кольца - к гиперполяризации. Из-за того, что она деполяризуется, когда включается свет, считается, что D-биполярная клетка имеет рецептивное поле с "on"-центром. Терминология "on" - и "off"-клеточных ответов будет активно использоваться нами далее при описании свойств рецептивных полей на следующих этапах зрительной системы. Важным принципом является то, что отдельные фоторецепторы могут одновременно находиться в центре одних рецептивных полей, контролируемых "on" - либо "off"-биполярной клеткой, в то же самое время являясь периферией в других рецептивных полях.
Горизонтальные клетки и ингибирование периферии
Ответы D - и Η-биполярных клеток при освещении периферии рецептивных зон опосредуются горизонтальными клетками. Каждая горизонтальная клетка получает информацию от большого количества фоторецепторов. Горизонтальные клетки, подобно Н-биполярным клеткам, отвечают на освещение фоторецепторов гиперполяризацией (из-за того, что при освещении уменьшается выделение фоторецепторами глутамата). Другой особенностью горизонтальных клеток является то, что они связаны друг с другом посредством электрических синапсов. Краситель люцеферовый желтый (Lucifer yellow), введенный в одну горизонтальную клетку, распространяется через щелевые контакты в другие нейроны. Таким образом, каждая горизонтальная клетка получает влияние с большой зоны сетчатки, иннервируемой ею и ее ближайшими соседями.
Горизонтальные клетки образуют с биполярными и фоторецепторными клетками тормозные синаптические соединения, в которых медиатором служит ГАМК. Таким образом, деполяризации фоторецепторов в темноте противостоит тормозное влияние горизонтальных клеток. Освещение фоторецепторов приводит к гиперполяризации горизонтальных клеток, и, следовательно, к уменьшению выделения ими ГАМК. Получается, что гиперполяризации фоторецепторов рассеянным светом противостоит соответствующее уменьшение секреции ГАМК в синапсах между фоторецептором и горизонтальной клеткой.
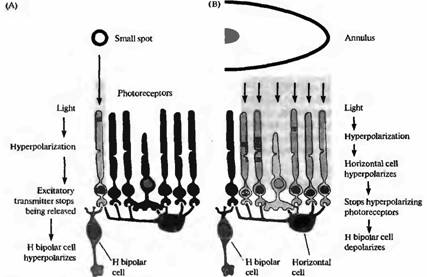
Рис. 5. Соединения между фоторецепторами биполярными и горизонтальными клетками. (А) Свет, падая на отдельную фоторецепторную клетку, вызывает ее гиперполяризацию, результатом чего является прекращение выделения глутамата В результате H биполярная клетка, как показано на рис. 19.18, гиперполяризуется, уменьшая свое возбуждение. (В) Свет, падающий на соседние области в форме кольца, также приводит к тому, что уменьшается высвобождение глутамата фоторецепторами. В результате происходит гиперполяризация горизонтальной клетки, которая предотвращает высвобождение ею тормозного медиатора ГАМК на фоторецептор. Следовательно, фоторецептор, который соединен с Η-биполярный клеткой, деполяризуется (из-за уменьшения тормозного влияния), что вновь приводит к высвобождению медиатора и деполяризации биполярной клетки. При освещении рассеянным светом процессы гиперполяризации и деполяризации гасят друг друга. Таким образом, горизонтальные клетки играют ведущие роли для обеспечения характерных свойств рецептивных полей биполярных клеток.
Было высказано предположение, что определенную роль здесь играет оксид азота, синтезируемый как фоторецепторами, так и горизонтальными клетками, которые также вносят вклад в угнетение высвобождения глутамата фоторецепторами. В итоге отрицательная обратная связь между фоторецепторами и горизонтальными клетками происходит по следующей схеме: свет. > гиперполяризация фоторецепторов > гиперполяризация горизонтальных клеток > деполяризация фоторецепторов.
Соединения клеток в случае центрального "on" - и периферического "off"-ответов биполярных клеток схематично показаны на рис.5. Для простоты центральная зона представлена всего одним фоторецептором, а периферия - несколькими соседними фоторецепторами, связанными с одной горизонтальной клеткой.
Сигнал в ответ на освещение центрального фоторецептора очевиден (рис.5А). Активация фоторецептора приводит к его гиперполяризации и, следовательно, к уменьшению высвобождения глутамата, что вызывает гиперполяризацию биполярной клетки. Горизонтальная клетка также получает стимулы к гиперполяризации, однако, так как они поступают только с одного фоторецептора, они имеют небольшой эффект, проявляющийся только в виде отрицательной обратной связи на центральный фоторецептор.
Ответы на освещение периферии (рис.5В) вовлекают дополнительные механизмы. Горизонтальная клетка, получающая сигналы от нескольких фоторецепторов вокруг центральной зоны, при освещении гиперполяризуется. Гиперполяризация уменьшает высвобождение ГАМК горизонтальными клетками, что приводит к уменьшению тормозного влияния на фоторецепторы и к их деполяризации. Этот эффект деполяризации проявляется минимально по отношению к фоторецепторам периферии, так как они сами по себе гиперполяризованы освещением. Однако центральный фоторецептор не находится под освещением, и поэтому единственным влиянием на него является уменьшение ингибирования со стороны горизонтальной клетки. В результате этого центральная клетка деполяризуется, увеличивается высвобождение ею глутамата, что все вместе приводит к деполяризации Η-биполярной клетки. В обзорах подробно описываются свойства терминалей фоторецепторов, терминалей биполярных клеток, а также синапсы обратной связи между горизонтальными и биполярными клетками.