Влияние цитокинина на рост и развитие проростков пшеницы в зависимости от условий минерального питания
В настоящее время известно четыре группы фитогормонов:
ауксины, гибберелины, цитокинины и ингибиторы роста. Все они могут активизировать, но могут и тормозить функциональную активность клеток. Общим условием для действия любого фитогормона является наличие в клетках специфичных рецепторов. Все фитогормоны вызывают у компетентных клеток сравнительно быстрые физиологические реакции, связанные, очевидно, с мембранами и более медленные изменения, зависящие от синтеза белков и нуклеиновых кислот ( Полевой В., 1982 ).
Помимо естественных фитогормонов, получено большое количество синтетических аналогов этих природных соединений, которые часто обладают высокой физиологической активностью. Строго говоря, эти вещества не могут отнесены к фитогормонам, так как не образуются в растениях, однако многие из них по активности не уступают фитогормонам или даже превосходят их ( Муромцев Г.С., 1973 ).
2.2 Общая характетистика цитокининов
а) Химическая структура
Цитокининами называется один из типов фитогормонов, обладающих определенной совокупностью биологической активности, которая весьма разнообразна и проявляется при регуляции роста, органообразования, процессов старения и покоя.
Цитокинины принимают участие наряду с другими фитогормонами в регуляции самых разнообразных физиологических процессов в растении. Для них, как и для других фитогормонов, характерна полифункциональность. Кроме того, действие цитокининов появляется в неразрывной связи с действием других фитогормонов и природных ингибиторов.
В химическом отношении природные цитокинины и их синтетические заменители представляют собой производные
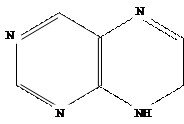 6-аминопурина с заместителем в аминогруппе при шестом атоме углерода пуринового кольца.
6-аминопурина с заместителем в аминогруппе при шестом атоме углерода пуринового кольца.
Пурин
Цитокинины пуринового ряда слаборастворимы в воде, но хорошо растворимы в этаноле, этиловом эфире, ряде щелочей и кислот. Цитокинины устойчивы к нагреванию, автоклавированию, действию щелочей и кислот.
Однако точная химическая характеристика цитокининов, как производных 6-аминопурина, затрудняется тем, что такими же свойствами обладает также дифенилмочевина и ее многочисленные производные. Очевидно, вопрос о химической характеристике цитокининов получит окончательное разъяснение только после установления молекулярного механизма их действия.
б) История открытия
Цитокинины были открыты в 1955 г. Скугом и Миллером с сотрудниками в Висконсинском университете в США. Их обнаружению помог так называемый “дефектный объект”, которым явился каллюс сердцевины стебля табака. Он образовался на кусочках сердцевины стебля в условиях стерильной культуры, но быстро прекращал рост в связи с исчерпанием какого-то фактора, исходно присутствовавшего в тканях стебля. Ввиду того, что вернуть активный рост каллюса удавалось добавлением к питательной смеси, содержащей индолилуксусную кислоту ( ИУК ), дрожжевого экстракта, а так же кокосового молока, Скугом и сотрудниками была предпринята попытка выделить недостающие для роста каллюса вещество из дрожжевого экстракта. При этом выяснилось, что активное вещество обладает свойствами пурина.
Вместе с тем проверка природных пуриновых оснований так же как гидрометода РНК и ДНК показала, что они не способны обеспечить рост стеблевого каллюса табака. Неожиданно активным оказался эфирный экстракт из старого, давно хранившегося препарата ДНК. Впоследствии выяснилось, что активное вещество образуется в любом препарате ДНК при ее деградации путем автоклавирования в кислой среде. Это вещество было выделено в виде кристаллов и идентифицировано химически. Оно оказалось 6- фурфуриламинопурином.
6-фурфуриламинопурин
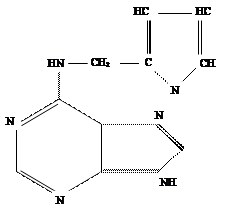 |
Указанное соединение образуется при автоклавировании ДНК в кислой среде из дезоксиаденозина. Поскольку 6-фурфуриламино-пурин в присутствии ИУК обеспечивал клеточное деление в изолированной сердцевине ткани стебля табака, он был назван кинетином ( от слова “кинез” ).
Принцип химического синтеза производных пурина с замещением в аминогруппе у шестого атома углерода пуринового кольца был известен ранее, поэтому вслед за химической идентификацией кинетина последовал его синтез. Затем было синтезировано много других активных соединений, которые отличались от кинетина характером заместителя в аминогруппе при шестом атоме углерода. Все эти соединения, включая кинетин, были объединены под общим названием кинины. Это название было распространено в литературе до 1965 г. Однако ввиду того, что в физиологии животных термин “кинины” еще ранее был применен для совершенно иных соединений - биологически активных
полипептидов, Скуг, Стронг и Миллер предложили заменить название кинины на цитокинины. В настоящее время термин цитокинины стал общепризнанным.
2.3 Физиологическая роль цитокининов
а) Стимуляция деления клеток
Цитокинины были открыты как вещества, необходимые для деления клеток у стеблевого каллюса табака и изолированной сердцевинной ткани стебля табака (Мi11еr, 1956; Skoog, Мil1еr, 1957). Эти ткани не способны к синтезу цитокининов, а также ауксинов, что позволило выявить многие существенные стороны действия данных фитогормонов на ростовые процессы. Так, в работах Скуга с сотрудниками было показано, что у изолированной сердцевинной ткани стебля табака в стерильной культуре одна ИУК несколько усиливала синтез ДНК, вызывала в отдельных клетках митозы, но не индуцировала клеточных делений. Точно так же один кинетин не вызывал деления клеток. В его присутствии не происходили митозы, а синтез ДНК он стимулировал в меньшей степени, чем ИУК. Только совместное действие ауксинов и кинетина значительно активировало синтез ДНК, вызывало митозы и индуцировало деление, клеток в изолированной сердцевине стебля табака (Dаs еt а1., 1956, 1958).
В последующем были предприняты попытки разграничить во времени действие ИУК и кинетина в индукции клеточных делений у этой ткани (Nitsch, 1968; Дмитриева, 1972). Полученные данные не имеют пока однозначной интерпретации, но позволяют предполагать, что начальные стадии процесса индуцируются одним ауксином, тогда как в последующем необходимы оба гормона ( Дмитриева, 1972 ).
Таким образом, было установлено, что регуляция деления клеток в изолированной сердцевинной ткани стебля табака осуществляется не одним гормоном - гипотетическим индуктором клеточных делений, а одновременным действием двух гормонов - цитокинина и ауксина. При этом для каждого фитогормона был показан свой предел концентраций, в которых он при наличии в среде другого гормона стимулирует данный процесс. Например, кинетин в присутствии в питательной среде 2 мг/л ИУК стимулировал деление клеток сердцевинной ткани стебля табака, начиная с концентрации 0,005 мг/л, но максимальное его действие достигалось в пределах концентраций от 0,02 до 0,5 мг/л. Увеличение концентрации кинетина до 2 мг/л приводило к угнетению процесса. Стимулирующее действие ИУК при одном и том же содержании в среде кинетина нарастало с увеличением ее концентрации до 1,8 мг/л. Максимальную интенсивность деления клеток вызывало сочетание оптимальных концентраций в среде обоих фитогормонов ( Scoog , Miller, 1957 ) .