Генетика и биометрия
Эпистатическое взаимодействие неаллельных генов в определенном смысле противоположно комплементарному действию генов. Сущность эпистаза сводится к подавлению проявления генов одной аллельной пары генами другой. Гены, подавляющие действие других неаллельных генов, называются супрессорами или подавителями. Они могут быть как доминантными, так и рецессивными, например А - В- или bbA - . Наследование окраски у свиней демонстрирует доминантный Эпистаз. При скрещивании черных и белых свиней из разных пород в F1 появляются белые потомки. Их скрещивание между собой приводит к появлению белых (12/16), черных (3/16) и красных (1/16) поросят. Все белые поросята имеют минимум один доминантный генподавитель I. Черные поросята гомозиготны по рецессивному аллелю i, не препятствующему формированию окраски, и несут доминантный аллель Е, детерминирующий образование черного пигмента. Красные поросята (eeii) лишены доминантного геноподавителя I и доминантного гена, определяющего черную окраску.
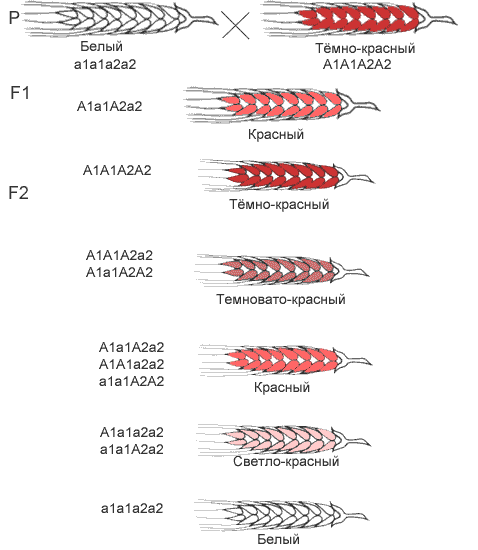
В некоторых случаях установлено, что проявление конкретного признака зависит от количества доминантных генов, вносящих вклад в его развитие. Например, при скрещивании краснозерных пшениц с белозерными было установлено, что растения с генотипом А1А1А2А2 имеют красные зерна, растения а1а1а2а2 – белые зерна, растения с тремя доминантными генами – красноватую окраску, а растения с двумя и одним геном – более бледную окраску. Таким образом, накопление определенных аллелей в генотипе может вести к изменению выраженности признаков.
2. Теория Ф. Жакоба и Ж- Моно о регуляции синтеза и-РНК и белков
Механизм регуляции генетического кода был открыт французскими учеными Ф. Жакобом и Ж. Моно в 1961 г. на бактериях. E. coli и получил название механизма индукции-репрессии. Было установлено, что синтез соответствующих белков – ферментов – индуцируется веществом, служащим субстратом и необходимым для нормальной жизнедеятельности клетки. Так, например, для нормальной жизнедеятельности E. coli необходим молочный сахар (лактоза), и в ее геноме содержаться гены, контролирующие синтез ферментов, гидролизующих лактозу до простых соединений. Если среда, в которой находятся бактерии, лактозы не содержит, эти гены пребывают в репрессированном состоянии и не функционируют. Внесенная в среду лактоза будет тем индуктором, который включает в работу длинные гены, и в клетке начинается синтез ферментов, гидролизующих лактозу до более простых соединений. После удаления лактозы из среды синтез этих ферментов прекращается. Роль репрессора может выполнять и вещество, синтезируемое в клетке, если содержание его превышает норму. Например, если синтезируются нуклеотиды аминокислоты и другие вещества и содержание их превышает количество, необходимое данной клетке, каждое из них может быть репрессором и подавлять работу генов, синтезирующих ферменты, необходимые для данного биохимического процесса.
Механизм индукции-репрессии обеспечивает включение в работу тех генов, которые синтезируют необходимые на данном этапе жизнедеятельности клетки ферменты. Работа генов прекращается, когда деградируемый данными ферментами субстрат израсходован или когда синтезируемое данными ферментами вещество находится в избытке. У высших организмов процесс регуляции работы генов осуществляется более сложно: у животных важную роль в этом процессе играют гормоны, клеточные мембраны; у растений - условия внешней среды, в том числе и окружающие клетки.
Раскрытие механизма регуляции генетического кода показало сложное строение локализованного в молекуле ДНК генетического аппарата. Гены, непосредственно кодирующие синтез соответствующих ферментов, называют структурными генами. Они входят в состав оперона, работу которого регулирует ген-регулятор. Как правило, структурные гены в опероне находятся в состоянии репрессии. Ген-регулятор расположен на особом участке молекулы ДНК и кодирует синтез специального белка, называемого репрессором. Работой структурных генов управляют находящиеся в опероне гены, не имеющие кодирующих функций. Их называют акцепторными генами. Система акцепторных и структурных генов образует единицу генетической регуляции, или оперон.
Акцепторные гены служат местом прикрепления различных белков, регулирующих работу структурных генов. Если лактоза, проникая в клетку, блокирует белки, кодируемые геном-регулятором, то они теряют способность присоединяться к гену-оператору. Ген-оператор переходит в активное состояние и включает в работу структурные гены. РНК-полимераза с помощью Сар-белка присоединяется к промотору и, продвигаясь вдоль оперона, синтезирует про-м РНК. При транскрипции мРНК считывает генетическую информацию со всех трех структурных генов в одном опероне. При трансляции на рибосоме происходит синтез трех разных полипептидных цепей в соответствии с содержащимися в мРНК кодами – последовательностями нуклеотидов, обеспечивающих инициацию и терминацию трансляции каждой цепи.
Тип регуляции работы генов, рассмотренной на примере лактозного оперона, называется негативной индукцией синтеза белка. Другим типом регуляции работы генов служит негативная репрессия, изученная у E.coli на примере trp –оперона, контролирующего синтез аминокислоты триптофана. Этот оперон состоит из 6700 пар нуклеотидов и содержит пять структурных генов, ген-оператор и два промотора. Ген-регулятор обеспечивает постоянный синтез регулярного белка, который не влияет на работу trp-оперона. При избытке в клетке триптофана последний соединяется с регуляторным белком и изменяет его таким образом, что он связывается с опероном и репрессирует синтез соответствующей мРНК.
Известна также и так называемая позитивная индукция, когда белковый продукт гена-регулятора активирует работу оперона, то есть выступает в роли не репрессора, а активатора. Деление это условно, и строение акцепторной части оперона, действие гена-регулятора у прокариотов весьма разнообразны.
Число структурных генов в опероне у прокариотов колеблется от одного до двенадцати; оперон может иметь либо один, либо два промотора и терминатора. Все структурные гены, локализованные в одном опероне, как правило, контролируют систему ферментов, обеспечивающих одну цепь биохимических реакций. Несомненно, что в клетке существуют системы, согласующие регуляцию работы нескольких оперонов.
В связи с особенностями организации отдельных генов эукариот и генома в целом регуляция генной активности у них характеризуется некоторыми отличиями по сравнению с прокариотами.
У эукариот не обнаружено организации генов по типу оперона. Установлено, что функционирование генов, несомненно, подчиняется регуляторным воздействием, однако регуляция транскрипции у эукариот является комбинационной, то есть активность каждого гена регулируется большим спектром генов-регуляторов.
У эукариотических генов имеется несколько областей, которые узнаются разными белками-регуляторами. Одна из них расположена недалеко от промотора и включает около ста пар нуклеотидов, в том числе ТАТА-блок. Установлено, что для успешного присоединения РНК-полимеразы к промотору необходимо предварительное соединение с ТАТА-блоком особого белка – фактора транскрипции – с образованием стабильного транскрипционного комплекса. Именно этот комплекс ДНК с белком узнает РНК-полимеразой. Последовательности нуклеотидов, примыкающих к ТАТА-блоку, формируют требуемый для транскрипции элемент, расположенный перед промотором.