Гормоны
Животный организм адаптируется к характеру пищевого режима, к нервной или мышечной нагрузке с помощью сложного комплекса координирующих механизмов. Так, контроль течения различных реакций углеводного и липидного обменов осуществляется на уровне клетки концентрациями соответствующих субстратов и ферментов, а также степенью накопления продуктов той или иной реакции. Эти контролирующие механизмы относятся к механизмам саморегуляции и реализуются как в одноклеточных, так и в многоклеточных организмах. У последних регуляция утилизации углеводов и жиров может происходить на уровне межклеточных взаимодействий. В частности, оба вида обмена реципрокно взаимоконтролируются: НЭЖК в мышцах тормозят распад глюкозы, продукты же распада глюкозы в жировой ткани тормозят образование НЭЖК. У наиболее высокоорганизованных животных появляется особый межклеточный механизм регуляции межуточного обмена, определяемый возникновением в процессе эволюции эндокринной системы, имеющей первостепенное значение в контроле метаболических процессов целого организма.
Среди гормонов, участвующих в регуляции жирового и углеводного обменов у позвоночных, центральное место занимают следующие: гормоны желудочно-кишечного тракта, контролирующие переваривание пищи и всасывание продуктов пищеварения в кровь; инсулин и глюкагон — специфические регуляторы межуточного обмена углеводов и липидов; СТГ и функционально связанные с ним «соматомедины» и СИФ, глюкокортикоиды, АКТГ и адреналин — факторы неспецифической адаптациии. Следует отметить, что многие названные гормоны принимают также непосредственное участие и в регуляции белкового обмена (см. гл. 9). Скорость секреции упомянутых гормонов и реализация их эффектов на ткани взаимосвязаны.
Мы не можем специально останавливаться на функционировании гормональных факторов желудочно-кишечного тракта, секретируемых в нервно-гуморальную фазу сокоотделения. Их главные эффекты хорошо известны из курса общей физиологии человека и животных и, кроме того, о них уже достаточно полно упоминалось в гл. 3. Более подробно остановимся на эндокринной регуляции межуточного метаболизма углеводов и жиров.
Гормоны и регуляция межуточного углеводного обмена. Интегральным показателем баланса обмена углеводов в организме позвоночных является концентрация глюкозы в крови. Этот показатель стабилен и составляет у млекопитающих примерно 100 мг% (5 ммоль/л). Его отклонения в норме обычно не превышают ±30%. Уровень глюкозы в крови зависит, с одной стороны, от притока моносахарида в кровь преимущественно из кишечника, печени и почек и, с другой — от его оттока в работающие и депонирующие ткани (рис. 2).
Приток глюкозы из печени и почек определяется соотношением активностей гликогенфосфорилазной и гликогенсинтетазной реакции в печени, соотношением интенсивности распада глюкозы и интенсивности глюконеогенеза в печени и отчасти в почке. Поступление глюкозы в кровь прямо коррелирует с уровнями фосфорилазной реакции и процессов глюконеогенеза. Отток глюкозы из крови в ткани находится в прямой зависимости от скорости ее транспорта в мышечные, адипозные и лимфоидные клетки, мембраны которых создают барьер для проникновения в них глюкозы (напомним, что мембраны клеток печени, мозга и почек легко проницаемы для моносахарида); метаболической утилизации глюкозы, в свою очередь зависимой от проницаемости к ней мембран и от активности ключевых ферментов ее распада; превращения глюкозы в гликоген в печеночных клетках (Левин и др., 1955; Ньюсхолм, Рэндл, 1964; Фоа, 1972). Все эти процессы, сопряженные с транспортом и метаболизмом глюкозы, непосредственно контролируются комплексом гормональных факторов.
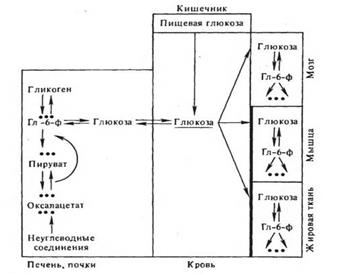
Рис.2. Пути поддержания динамического баланса глюкозы в крови Мембраны мышечных и адипозных клеток имеют "барьер" для транспорта глюкозы; Гл-б-ф — глюкозо-б-фосфат
Гормональные регуляторы углеводного обмена по действию на общее направление обмена и уровень гликемии могут быть условно разделены на два типа. Первый тип гормонов стимулирует утилизацию глюкозы тканями и ее депонирование в форме гликогена, но тормозит глюконеогенез, и, следовательно, вызывает снижение концентрации глюкозы в крови. Гормоном такого типа действия является инсулин. Второй тип гормонов стимулирует распад гликогена и глюконеогенез, а следовательно, вызывает повышение содержания глюкозы в крови. К гормонам этого типа относятся глюкагон (а также секретин и ВИП) и адреналин. Гормоны третьего типа стимулируют глюконеогенез в печени, тормозят утилизацию глюкозы различными клетками и, хотя усиливают образование гликогена гепатоцитами, в результате преобладания первых двух эффектов, как правило, также повышают уровень глюкозы в крови. К гормонам данного типа можно отнести глюкокортикоиды и СТГ — «соматомедины». Вместе с тем, обладая однонаправленным действием на процессы глюконеогенеза, синтеза гликогена и гликолиза, глюкокортикоиды и СТГ — «соматомедины» по-разному влияют на проницаемость мембран клеток мышечной и адипозной ткани к глюкозе.
По направленности действия на концентрацию глюкозы в крови инсулин является гипогликемическим гормоном (гормон «покоя и насыщения»), гормоны же второго и третьего типов — гипергликемическими (гормоны «стресса и голодания») (рис.3).
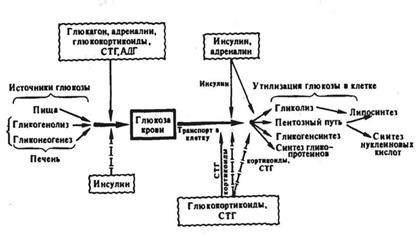
Рис 3. Гормональная регуляция углеводного гомеостаза: сплошными стрелками обозначена стимуляция эффекта, пунктирными — торможение
Инсулин можно назвать гормоном усвоения и депонирования углеводов. Одной из причин усиления утилизации глюкозы в тканях является стимуляция гликолиза. Она осуществляется, возможно, на уровне активации ключевых ферментов гликолиза гексокиназы, особенно одной из четырех известных ее изоформ — гексокиназы П, и глюкокиназы (Вебер, 1966; Ильин, 1966, 1968). По-видимому, определенную роль в стимуляции катаболизма глюкозы инсулином играет и ускорение пентозофосфатного пути на стадии глюкозо-6-фосфатдегидрогеназной реакции (Лейтес, Лаптева, 1967). Считается, что в стимуляции захвата глюкозы печенью при пищевой гипергликемии под влиянием инсулина важнейшую роль играет гормональная индукция специфического печеночного фермента глюкокиназы, избирательно фосфорилирующего глюкозу при высоких ее концентрациях.
Главная причина стимуляции утилизации глюкозы мышечными и жировыми клетками — прежде всего избирательное повышение проницаемости клеточных мембран к моносахариду (Лунсгаард, 1939; Левин, 1950). Таким путем достигается повышение концентрации субстратов для гексокиназной реакции и пентозофосфатного пути.
Усиление гликолиза под влиянием инсулина в скелетных мышцах и миокарде играет существенную роль в накоплении АТФ и обеспечении работоспособности мышечных клеток. В печени усиление гликолиза, по-видимому, важно не столько для повышения включения пирувата в систему тканевого дыхания, сколько для накопления ацетил-КоА и малонил-КоА как предшественников образования многоатомных жирных кислот, а следовательно, и три-глицеридов (Ньюсхолм, Старт, 1973). Образующийся в процессе гликолиза глицерофосфат также включается в синтез нейтрального жира. Кроме того, и в печени, и особенно в адипозной ткани для повышения уровня липогенеза из глюкозы существенную роль играет стимуляция гормоном глюкозо-б-фосфатдегидрогеназной реакции, приводящей к образованию НАДФН — восстанавливающего кофактора, необходимого для биосинтеза жирных кислот и глицерофосфата. При этом у млекопитающих только 3-5% всасываемой глюкозы превращается в печеночной гликоген, а более 30% накапливается в виде жира, откладываемого в депонирующих органах.