Деятельность центральной нервной системы
Тела нейронов, участвующих в образовании пирамидных трактов, лежат в двигательных зонах коры, а их аксоны образуют синапсы непосредственно с мотонейронами спинного мозга в том его сегменте, где эти нейроны выходят на периферию. В головном мозге нет никаких промежуточных синапсов, поэтому импульсы и последующие ответы на них по пути не задерживаются и не видоизменяются.
Главным экстрапирамидным трактом является ретикулоспинальный тракт, переключающий импульсы от ретикулярной формации, которая лежит в стволе мозга между таламусом и продолговатым мозгом. Из различных отделов головного мозга, контролирующих двигательную активность, импульсы поступают в определенные участки ретикулярной формации, где они модифицируются под воздействием импульсов, идущих от коры, и становятся либо возбуждающими, либо тормозными. Например, импульсы от мозжечка и премоторной зоны коры, управляющей координированными движениями, поступают в ту область ретикулярной формации, которая находится в продолговатом мозгу и посылает импульсы, стимулирующие тормозные мотонейроны. Последние подавляют активацию определенных мышц, что дает возможность осуществлять сложные координированные движения тела. Другие комбинации двигательных импульсов, напротив, стимулируют возбуждающие нейроны, и общее воздействие ретикулярной формации на двигательную активность оказывается возбуждающим.
Большинство волокон сенсорных нейронов на своем пути через таламус к коре отдает коллатерали (боковые ветви) в ретикулярную формацию, участвуя в образовании ретикулярной активирующей системы, которая тонизирует кору и участвует в пробуждении организма от сна. Недостаточная активность этой системы или ее разрушение приводит соответственно к глубокому сну или коме . Как полагают, многие вещества, вызывающие общий наркоз, оказывают свое действие, временно блокируя синаптическую передачу в этой системе. Предполагается так же, что ретикулярная активирующая система ответственна за возникновение и поддержание побуждений к действию и концентрации внимания.
Функции некоторых участков коры, в частности обширных передних областей - префронтальных зон коры головного мозга - остаются еще неясными. Эти области, а также ряд других участков мозга, называют немыми зонами коры головного мозга, так как при раздражении их электрическим током не возникает никаких ощущений или реакций. Предполагают, что эти зоны ответственны за наши индивидуальные особенности, или личность. Удаление этих зон или перезку проводящих путей, идущих от них к остальному мозгу (префронтальную лоботомию) применяли для снятия у больных острого возбуждения, но от этого пришлось отказаться из-за таких побочных эффектов, как снижение уровня сознания и интеллекта, способности к логическому мышлениию и способности к творчеству. Эти побочные эффекты косвенно указывают на функции, выполняемые префронтальными зонами.
Базальные ганглии расположены в основании конечного мозга и представляют собой важное подкорковое связующее звено между ассоциативными областями коры головного мозга и двигательными областями коры головного мозга. К базальным ганглиям относятся следующие структуры: полосатое ядро базальных ганглий (стриатум), состоящее из хвостатого ядра базальных ганглий и скорлупы базальных ганглий, бледный шар базальных ганглий (паллидум), подразделяющийся на внутренний и внешний отделы, черная субстанция базальных ганглий и субталамическое ядро базальных ганглий. В состав базальных ганглиев часто включают также ограду и реже - миндалину.
Рис.13
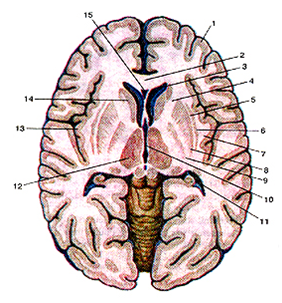
Базальные (подкорковые) узлы и внутренняя капсула на горизонтальном разрезеголовного мозга. Вид сверху.
1-кора большого мозга (плащ); 2-колено мозолистого тела; 3-пе-редний рог бокового желудочка; 4-внутренняя капсула; 5-каруж-ная капсула; 6-ограда; 7-самая наружная капсула; 8-скорлупа; 9-бледный шар; 10-III желудочек; 11-задний рог бокового желудочка; 12-thalamus (зрительный бугор); 13-корковое вещество (кора) островка; 14-головка хвостатого ядра; 15-полость прозрачной перегородки.
Базальные ядра облегчают запускаемые корой движения и подавляют лишние сопутствующие движения. Стриатум получает соматотопически организованные проекции почти от всех зон коры. Эти проекции организованы в виде параллельных путей, которые начинаются от лобных областей, постцентральных (соматосенсорных) областей, прецентральных (моторных) областей и теменно-височно-затылочных областей. В то же время прямое отношение к регуляции движений имеют только глутаматергические, возбуждающие проекции от моторной коры и соматосенсорной коры .
Нейроны стриатума посылают волокна к структурам, формирующим главный эфферентный путь базальных ядер - к сетчатой части черной субстанции и медиальному бледному шару. Эти структуры, в свою очередь, образуют тормозные ГАМК ергические синапсы на переднем и латеральном вентральных ядрах таламуса, которые направляют к коре возбуждающие глутаматергические волокна.
Таким образом, при торможении нейронов сетчатой части черной субстанции и медиального бледного шара возбуждающее влияние таламуса на кору усиливается - что и способствует облегчению запускаемого корой движения.
Напротив, при возбуждении нейронов сетчатой части черной субстанции и медиального бледного шара возбуждающее влияние таламуса на кору подавляется - что приводит к торможению ненужного движения.
Прямого выхода на спинной мозг базальные ядра не имеют. Прямой путь от стриатума к сетчатой части черной субстанции и медиальному бледному шару образован тормозными ГАМКергическими волокнами. Функция их состоит в усилении возбуждающего влияния ядер таламуса на те отделы моторной коры, которые отвечают за нужное движение.
Непрямой путь организован сложнее. Функция его заключается в подавлении возбуждающего влияния таламуса на другие отделы моторной коры. Первое звено этого пути - тормозные ГАМКергические проекции стриатума на латеральный бледный шар. Латеральный бледный шар посылает тормозные ГАМКергические волокна к субталамическому ядру. Выходы субталамического ядра представлены возбуждающими глутаматергическими волокнами - часть их возвращается к латеральному бледному шару, другие направляются к сетчатой части черной субстанции и медиальному бледному шару.
Таким образом, если активация прямого пути от стриатума усиливает возбуждение моторной коры, то активация непрямого пути - ослабляет.
Активность этих двух путей регулирует компактная часть черной субстанции, посылающая к стриатуму дофаминергические волокна. Эти волокна возбуждают прямой эфферентный путь стриатума (через нейроны с D1-рецепторами) и тормозят - непрямой (через нейроны с D2-рецепторами). Стриатум посылает также тормозные ГАМКергические волокна к компактной части черной субстанции, образуя тем самым с ней отрицательную обратную связь. Наконец, активность стриатума модулируют его холинергические вставочные нейроны - антагонисты дофаминергических нейронов.