Динамическое поведение мембранных систем и липидно-белковые взаимодействия
Введение
Все биологические структуры по своей природе динамичны, и при рассмотрении их функций необходимо учитывать подвижность компонентов, из которых эти структуры состоят. Это относится к ферментам, полинуклеотидам и, конечно, к мембранам. В жидкостно-мозаичной модели, в центре которой находится представление о подвижности мембранных компонентов, мембрана рассматривается как некое липидное море, в котором свободно плавают глобулярные белки. За последние два десятилетия было опубликовано множество работ, посвященных количественным и качественным аспектам динамики мембранных компонентов. Во многих из них использовались спектральные методы и спектроскопия магнитного резонанса. В результате была создана весьма полезная физическая картина мембран, иллюстрирующая способы перемещения мембранных белков и липидов и их взаимодействия. Кроме того, были разработаны специальные методики, направленные на изучение динамических свойств мембран. Многое в этой области остается неясным, но уже определены основные направления будущих исследований.
Главным побудительным мотивом в изучении мембранной динамики служит ее связь с биологическими функциями мембран. Необходимым условием протекания одних ферментативных процессов является свободная диффузия мембраносвязанных компонентов в плоскости бислоя, другие же процессы могут осуществляться лишь при ограниченной подвижности мембранных компонентов. Между шероховатым и гладким эндоплазматическими ретикулумами, комплексом Гольджи и плазматической мембраной происходит быстрый обмен различными веществами, и тем не менее их состав и функции различаются. Чтобы понять суть этих и многих других биологических феноменов, необходимо прежде всего выяснить фундаментальные аспекты динамических свойств мембран. Поперечная асимметрия в распределении липидов, а возможно, и пассивная проницаемость бислоев очевидным образом связаны со скоростью трансмембранного флип-флоп-переноса липидов. Биогенез мембран зависит от скорости обмена липидов между различными мембранами. Скорость ферментативных реакций, протекающих с участием мембраносвязанных компонентов, зависит от скорости латеральной диффузии компонентов мембран. Наконец, липидно-белковые взаимодействия зависят от скорости, с которой происходит обмен липидами между ближайшим окружением белков и остальным объемом мембраны.
Диапазон движений, происходящих в мембране, весьма широк: от молекулярных колебаний с частотой порядка 1014 с до трансмембранного флип-флоп-переноса липидов, характерное время которого может достигать несколько суток. На рис. 1 в общем виде представлены некоторые из этих процессов, а также указаны временные пределы чувствительности различных биофизических методов. Величины, приведенные на рисунке, различаются на 20 порядков, поэтому термины «быстрый» и «медленный» не могут адекватно характеризовать различные типы движений. Из рисунка видно также, что одни методы позволяют получить статичную картину мембраны, поскольку характерное время соответствующих движений больше, чем время измерений, в то время как другие методы дают усредненную по времени картину, поскольку время перемещения молекул гораздо меньше, чем время измерения.
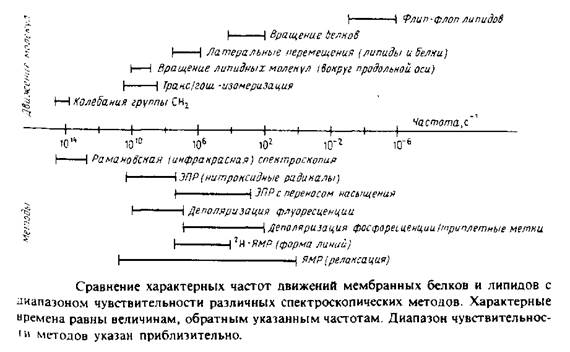
Мы рассмотрим два основных типа экспериментов. Первые основаны на использовании внутримембранных зондов для изучения текучести мембраны. Индикаторами физического состояния мембраны, а также характера липидно-белковых взаимодействий могут служить низкомолекулярные ЭПР-метки и флуоресцентные зонды.
Второй тип экспериментов направлен на прямое измерение латеральной диффузии мембранных белков или липидов и вращательной способности белков внутри бислоя. Исследовались также молекулярные взаимодействия в бислое, поскольку они влияют на динамику изучаемых молекул.
1. Некоторые простые модели движения мембранных компонентов
На рис. 2 представлены некоторые модели, использующиеся для анализа поступательного движения молекул внутри мембранного бислоя. Такие модели необходимы для интерпретации экспериментальных данных с точки зрения молекулярного движения.
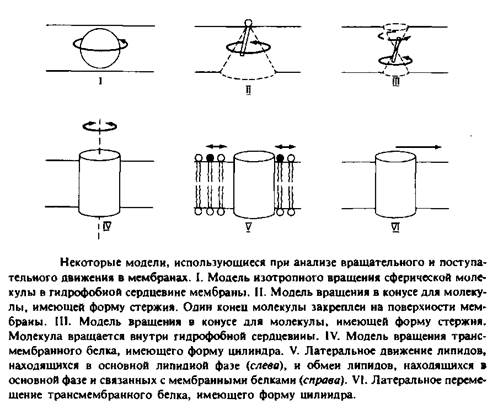
Модель изотропного вращения предполагает, что молекула вращается с одинаковой вероятностью во всех направлениях и выделенная ось вращения отсутствует. Подобное вращение будет совершать сферическая частица в непрерывной жидкой среде. Эта концепция применялась при анализе движения малых растворимых в мембране гидрофобных меток, таких, как TEMPO. Изотропная трехмерная диффузия сферической частицы рассматривается как случайное блуждание и характеризуется коэффициентом вращательной диффузии DBp. Этот параметр определяется как среднеквадратичное угловое отклонение или время релаксации.
2.Модель вращения в конусе описывает движение амфифильных меток, например производных жирных кислот. Эти молекулы можно представить в виде жестких стержней, один конец которых закреплен на поверхности мембраны. Их движение ограничивается конусом, который они описывают относительно некой оси, перпендикулярной плоскости мембраны.
3.Модель вращения в конусе используется в некоторых работах для описания движения ДФГ — гидрофобного флуоресцентного зонда, представляемого в виде жесткого стержня. В данном случае эта модель, постулирующая существование предпочтительной ориентации для ДФГ, является, безусловно, слишком упрощенной, поэтому были предложены альтернативные модели.
4. В простейшей модели вращения белков белковая молекула представляется в виде цилиндра, закрепленного в мембране и способного вращаться только вокруг оси, перпендикулярной плоскости мембраны. Такое вращение анизотропно. Время релаксации, характеризующее это движение, обычно представляется в виде 4>ц = \/. Знак «II» указывает на то, что вращение происходит
параллельно оси цилиндра.
5 и 6. Поступательное движение липидных и белковых молекул описывается как двумерная диффузия. Особый интерес представляет скорость обмена липидов, связанных с мембранными белками и находящихся в основном объеме мембраны. Изотропная двумерная диффузия характеризуется среднеквадратичным перемещением </2>, происходящим за время ![]()
Таблица 1. Некоторые метки, использующиеся для изучения динамики мембран
Для обычной жидкости, какой является, например, вода, текучесть определяется как величина, обратная вязкости — понятному и легко измеряемому физическому параметру. Вязкость характеризует трение, возникающее между соседними слоями жидкости, которые движутся с разными скоростями. Вязкость жидкости можно оценить, измерив скорость, с которой падает мраморный шарик в жидкости. В случае мембран термин «текучесть» обычно носит скорее качественный характер: имеется в виду сопротивление, которое оказывает мембрана различным типам перемещений в ней. Как правило, для измерения текучести наблюдают за движением спиновых или флуоресцентных зондов, включенных в мембрану. Зондами обычно являются небольшие молекулы, сравнимые по размерам с мембранными фосфолипидами. Некоторые из них представлены в табл. 5.1. Теоретические основы использования спектроскопических методов для изучения молекулярных движений описаны в последующих разделах. Поскольку эти методы позволяют измерять как скорость движения, так и сопротивление этому движению, сведения о динамике и молекулярной упорядоченности даются вместе. Укажем некоторые моменты, существенные для количественной интерпретации данных по движению зондов внутри мембран.