Клещи рода dermacentor koch. 1844 в Крыму
Голодные личинки живут в природе не более 20-60 сут и нимфы - 35-70 сут (Покровская, 1951, 1953, Daniel et al., 1980, Ламанова, 1966). Поэтому, из-за эфемерности неполовозрелых форм зимовка обоих видов возможна только на стадиях голодных имаго и напитавшихся самок, а голодные или напитавшиеся, но не перелинявшие к зиме нимфы поги-бают. Напротив, голодные имаго отличаются исключительной долговечностью. Во многом длительность жизни зависит от условий существования и в частности зимовки - в открытых и сухих стациях клещи живут меньше, чем в закрытых с повышенной влажностью. Продолжительность жизни взрослых клещей достигает трех летних сезонов с момента вылупления личинок (Адамович, 1961, Разумова, 1966; Балашов, 1998). При этом в пер-вую зимовку выживает до100% клещей, во вторую - около 80, во время третьей зимовки, все гибнут (Покровская 1958, Ламанова 1966). Вследствие этого активизирующаяся весной или осенью популяция представляет смесь из особей не менее трех поколений.
Время выхода зимующих имаго и их активизация весной во многом зависит не только от климатических факторов данного года, но и от конкретных микроклиматических условий мест зимовки. Повсеместно перезимовавшие имаго активизируются ранней весной со сходом снега и наступлением первых теплых дней.
В Крыму, в природных местообитаниях, успешно перезимовавшие клещи D. marginatus встречаются уже в теплые февральские "окна", а в марте отмечается их максимальная активизация (средне - многолетняя численность - 0.33 экз. на флаго/час) (Рис 2).
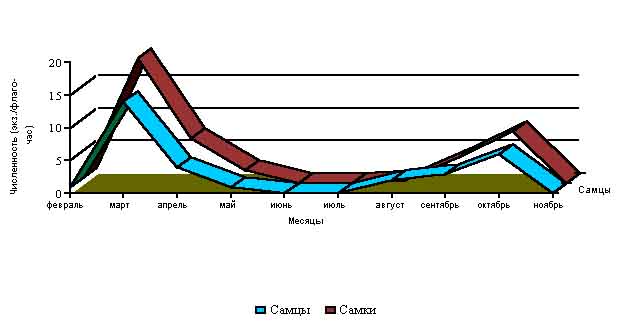 Рис. 2. Динамика численности клещей D.marginatus по месяцам (экз/флаго-час.)
Рис. 2. Динамика численности клещей D.marginatus по месяцам (экз/флаго-час.)
В местах с наиболее оптимальными условиями существования встречалось до 50-80 экз. на 1 флаго/час и больше. В апреле численность клещей снижается в три раза по сравнению с мартом, а в июне - еще в три раза по сравнению с маем. Причин такой динамики несколько. Во-первых, самки, нашедшие хозяев и успешно завершившие питание, уходят в подстилку, где начинают откладку яиц, после чего они погибают. Во-вторых, постоянно происходит естественная гибель особей от хищников, неблагоприятных климатических факторов и других причин. В-третьих, постепенно происходит повышение среднесуточных температур, сильно повышается сухость воздуха, что неблагоприятно влияет на активность клещей. В летние месяцы, характеризующиеся жаркой и сухой погодой (июнь - первая половина августа), оставшиеся в живых клещи покидают растительность и укрываются от неблагоприятного воздействия высоких температур, низкой влажности воздуха и сильной инсоляции в подстилке, впадая в летнюю диапаузу. После снижения среднесуточных температур и выпадения дождей, характерных для конца августа, происходит активизация клещей, что выражается в постепенном нарастании их численности вплоть до середины октября. Хотя суммарная численность имаго в популяциях в данный период намного выше таковой чем весной, однако большинство клещей нового поколения не активизируются и не покидают подстилку. С конца октября и в ноябре, когда температура становится отрицательной, клещи снова покидают растительность и укрываются в расщелинах почвы, подстилке и других укромных местах, где и зимуют. Отдельные особи клещей, в связи с нередкими для Крыма теплыми и погожими "окнами", которые случаются в ноябре-декабре, могут активизироваться и встречаться в активном состоянии на остатках травянистых растений и кустарниках.
Динамика численности D. reticulatus аналогична предыдущему виду (см табл. ). Только в связи с тем, что в верхнем поясе гор весной теплая погода устанавливается позже, соответственно и сроки активности клещей смещены на более поздние сроки. Так активизация клещей после зимовки происходит в марте, достигая своего пика в апреле, а с мая снижается, падая почти до нуля. Осенняя динамика активности клещей D. reticulatus примерно соответствует таковой клещей D. marginatus. Осенние волны заклещевления, как пра-вило, бывают менее выраженными, чем весенние. Отличительной особенностью клещей D. marginatus является их способность зимо-вать на хозяине, в то время как зимующих D. reticulatus на животных не обнаружено (Резник, 1970). Сборы, сделанные в холодное время года, мо-гут быть довольно обильными и содержать напившихся самок (зимовать остаются самки, присо-савшиеся поздно осенью) (Галузо, 1948).
Близкие данные по фенологии клещей отмечены и в других районах ареала (Адамович, 1961, Галузо, 1948, Разумова 1962, Ганиев, 1956). Однако иногда отмечаемое исследователями превышение осеннего пика численности над весенним (Сапегина, 1965, Богданов и др., 1964), в Крыму не наблюдалось. Длитель-ность обеих волн сезонной активности более или менее одинакова и продол-жается около двух месяцев (Логиновский, l966).
Поведенческая и репродуктивная диапаузы регу-лируются длиной светового дня. Для D. marginatus характерна короткодневная фотопериодическая реакция с критическим фотопериодом в 13 ч (Бе-лозеров, 1976). Экологическое значение обеих форм диапаузы однозначно и на-правлено на предотвращение откладки яиц в конце лета и осенью и гибели не достигнувших имагинальной фазы особей зимой. Самки, напитавшиеся с середины лета, и осенью находятся в состоянии репро-дуктивной диапаузы и приступают к яйцекладке только после зимовки вместе с особями весеннего насыщения В июне-июле клещи становятся неактивными и прекращают нападать на хозяев, хотя и могут оставаться на растительности. В сезонной регуляции главная роль принадлежит имагинальной диапаузе (Балашов, 1998).
Суточная активность
Половозрелые особи клещей р. Dermacentor суточных миграций практически не совершают, оставаясь на выбранных растениях постоянно. Это подтверждают специальные наблюдения, проведенные на Ай-Петринской яйле во второй декаде сентября. Клещи оставались на растительности в позе пассивного ожидания в течение всех суток, несмотря на резкие перепады температуры. Они одинаково активно цеплялись на флаг днем, когда температура воздуха достигла +150С; поздно вечером, после наступления сумерек при 5-70С тепла и рано утром, при падении температуры воздуха до 30С ниже нуля.
Сезонная регуляция одногодичного цикла обеспечивается существованием летней неативности у голодных и диапаузы у напитавшихся самок. Впадение клещей этих видов в состояние летней неактивности и возникновение несколько позднее потенциальной способности к диапаузе у напитавшихся самок служит надежной биологической адаптацией, предотвращающей появление в конце лета и осенью не способных к зимовке неполовозрелых фаз развития (Фильчагов, Лебедева, 1988).
Как показали наблюдения этих же авторов, прокормители собирают 10-20 активизировавшихся в агрегациях личинок. Разброс напитавшихся особей составляет около 1 га. Единичные заносы отмечены на расстояние до 300 м. Однако в отдельных случаях возможен занос птицами или более подвижными животными (например зайцем-русаком) и на более дальние расстояния.
Индивидуальные различия массы взрослых клещей (как и их размеры), зависят в основном от количества крови поглощенной на предимагинальных стадиях (в большей мере на нимфальной, в меньшей - на личиночной) и длительности вынужденного голодания в ожидании прокормителя. В третьей декаде марта 1988 года, в трех различных биотопах нами были собраны клещи, вышедшие с мест зимовки. Сравнение полученных данных по индивидуальному взвешиванию живых клещей показало, что масса отдельных особей самцов колебалась от nmin=3,5 мг до nmax =18 мг, при средней массе самцов в трех выборках: 8,4+0,4; 9,4+0,9; 9,4+1,4 мг. Масса самок изменялась от nmin=3,0 мг до nmax =14,0 мг, при средней массе самок в трех выборках: 8,1+0,2; 7,5+0,3; 8,4+0,4 мг. Во всех трех локальных популяциях самцы достоверно крупнее самок и эта разница (по средним показателям) колеблется от 4,1% до 19,9%, что связано с наличием у самцов сильно хитинизированного покрова. Длина тела имаго в голодном состоянии колебалась в пределах 2-8,5 мм.