Меристемы или образовательные ткани
Первичные меристемы обладают меристематической активностью, т. е. способны к делению изначально. В ряде случаев способность к активному делению может вновь возникнуть и у клеток, уже почти утративших это свойство. Такие «вновь» возникшие меристемы называют вторичными.
В теле растения меристемы занимают различное положение, что позволяет их классифицировать. По положению в растении выделяют верхушечные, или апикальные (от лат. «апекс» — верхушка), боковые, или латеральные (от лат. «латус» — бок), и интеркалярные меристемы.
Апикальные меристемы располагаются на верхушках осевых органов растения и обеспечивают рост тела в длину, а латеральные — преимущественно рост в толщину. Каждый побег и корень, а также зародышевый корешок, почечка зародыша имеют апикальную меристему. Апикальные меристемы первичны и образуют конусы нарастания корня и побега (рис. 1).
Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов, образуя своего рода цилиндры, на поперечных срезах имеющие вид колец. Часть из них относится к первичным. Первичными меристемами являются прокамбий и перицикл, вторичными — камбий и феллоген.
Интеркалярные, или вставочные, меристемы чаще первичны и сохраняются в виде отдельных участков в зонах активного роста (например, у оснований междоузлии, в основаниях черешков листьев).
Существуют также раневые меристемы. Они образуются в местах повреждения тканей и органов и дают начало каллюсу — особой ткани, состоящей из однородных паренхимных клеток, прикрывающие место поражения Каллюсо-образовательная способность растений используется в практике садоводства при размножении их черенками и прививками. Чем интенсивнее каллюсообразование, тем больше гарантия срастания подвоя с привоем и укоренения черенков. Образование каллюса— необходимое условие культуры тканей растения на искусственных средах.
Клетки апикальных меристем более или менее изодиаметричны по размерам и многогранны по форме. Межклетников между ними нет, оболочки тонкие, содержат мало целлюлозы. Полость клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение. Вакуоли многочисленные, мелкие, но под световым микроскопом обычно не заметны. Эргастические вещества, как правило, отсутствуют. Пластид и митохондрий мало и они мелки.
Клетки боковых меристем различны по величине и форме. Они примерно соответствуют клеткам тех постоянных тканей, которые из них в дальнейшем возникают. Так, в камбии встречаются как паренхимные, так и прозенхимные инициали. Из паренхимных инициалей образуется паренхима проводящих тканей, а из прозенхимных — проводящие элементы.
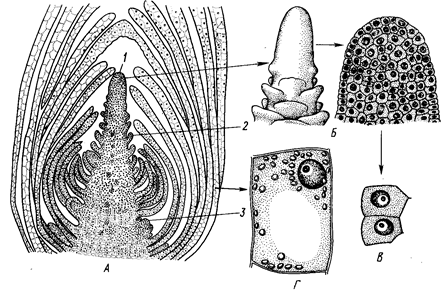
Рис. 1. Верхушечная меристема побега элодеи. А — продольный срез; 5 — конус нарастания (внешний вид и продольный срез); В — клетки первичной меристемы;
Г — паренхимная клетка сформировавшегося листа:
1 ~ конус нарастания, 2 — зачаток листа, 3 — бугорок пазушной почки
Зоны корня. Особенности морфологического строения каждой зоны в связи с выполняемыми функциями. Рост корня в длину и втолщину.
Наряду с побегом корень - основной орган высшего растения, в типичном случае выполняющий функцию минерального и водного питания. Другая важная функция корня, тесно связана с основной, - закрепление, растения в почве.
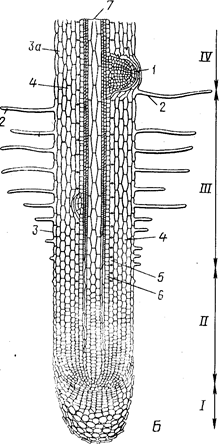
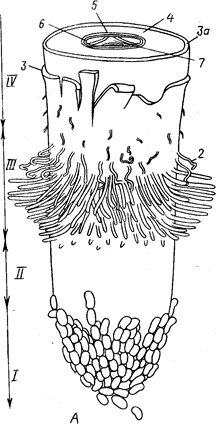
Различные части корня выполняют неодинаковые функции и характеризуются определенными морфологическими особенностями. Эти части получили название зон (рис. 2). Кончик корня снаружи всегда прикрыт корневым чехликом, защищающим апикальную меристему. Клетки корневого чехлика продуцируют слизь, покрывающую поверхность корня. Благодаря слизи снижается трение о почву, и ее частицы легко прилипают к корневым окончаниям и корневым волоскам.
Корневой чехлик состоит из живых паренхимных клеток, возникающих у большинства однодольных из особой меристемы, получившей название калиптрогена, а у двудольных и голосеменных — из верхушечной меристемы кончика корня. Водные растения корневого чехлика обычно не имеют.
Под чехликом располагается зона деления, представленная меристематической верхушкой корня, его апексом. В результате активности апикальной меристемы формируются все прочие зоны и ткани корня.
Делящиеся клетки сосредоточены в зоне деления, имеющей размеры около 1 мм. Эта часть корня заметно отличается от прочих зон своей желтоватой окраской. Вслед за зоной деления располагается зона растяжения (роста). Она также невелика по протяженности (несколько миллиметров), выделяется светлой окраской и как бы прозрачна. Клетки зоны роста практически не делятся, но способны растягиваться в продольном направлении, проталкивая корневое окончание в глубь почвы. Они характеризуются высоким тургором, что способствует активному раздвиганию частиц почвы. В пределах зоны роста происходит дифференциация первичных проводящих тканей.
Окончание зоны роста заметно по появлению на эпиблеме многочисленных корневых волосков. Корневые волоски располагаются в зоне—всасывания, функция которой понятна из ее названия. На корне она занимает участок от нескольких миллиметров до нескольких сантиметров. В отличие от зоны роста участки этой зоны уже не смещаются относительно частиц почвы. Основную массу воды и растворов солей молодые корни усваивают в зоне всасывания с помощью корневых волосков.
Корневые волоски появляются в виде небольших сосочков — выростов клеток эпиблемы. Рост волоска осуществляется у его верхушки. Оболочка корневого волоска растягивается быстро. По прошествии определенного времени корневой волосок отмирает. Продолжительность его жизни не превышает 10—20 дней.
Выше зоны всасывания, там, где исчезают корневые волоски, начинается зона проведения. Строение этой зоны на разных ее участках неодинаково. По этой части корня вода и растворы солей, поглощенные корневыми волосками, транспортируются в вышележащие отделы растения. Участки зоны проведения занимают фиксированное положение относительно участков почвы, не смещаясь относительно их. Несмотря на фиксированное положение в пространстве конкретных участков зон поглощения и проведения, эти зоны сдвигаются вследствие верхушечного роста. В результате этого всасывающий аппарат постоянно перемещается в почве.
В пределах одной корневой системы существуют корни, выполняющие разные функции. У большинства растений различаются ростовые и сосущие окончания. Ростовые окончания долговечнее, относительно мощные, быстро удлиняются и продвигаются в глубь почвы. Сосущие окончания недолговечны, возникают в большом числе на вростовых корнях
и удлиняются медленно. У деревьев и кустарников выделяют видоизмененные скелетные и полускелетные корни, на которых возникают недолговечные корневые мочки, несущие множество сосущих окончаний.
|
|
|
|