Механизмы дыхания растений
Кислород в пирокатеховой кислоте оказался меченым. Таким образом, в точном эксперименте было доказано, что молекулярный кислород действительно может включаться в органические соединения.
В настоящее время известно, что путь включения кислорода в органические соединения в соответствии с перекисной теорией биологического окисления Баха и Энглера не имеет отношения к дыханию, однако работы этих исследователей сыграли большую роль в изучении химизма дыхания, заложив основы современного понимания механизмов активации кислорода.
Какую же роль играет кислород в процессах дыхания? В 1921 г. немецкий биохимик О. Г. Варбург, изучая влияние ингибиторов на дыхание различных объектов, обнаружил, что поглощение кислорода резко ингибируется оксидом углерода и синильной кислотой, взаимодействующими в клетках с железосодержащим веществом порфириновой природы (с цитохромоксидазой). Английский биохимик Д. Кейлин в 1925 г. окончательно доказал присутствие в клетках цитохромоксидазы, ускоряющей поглощение ими кислорода, и открыл другие цитохромы. Затем цитохромы были обнаружены у всех аэробов и было показано, что у этих организмов на завершающем этапе процесса дыхания осуществляется перенос на кислород электронов и протонов, в результате чего образуется Н20 (или Н202).
Органические соединения могут окисляться и другим способом благодаря отнятию водорода. Еще Бах, сделав упор на перекисную теорию, выдвинул и вторую гипотезу, согласно которой биологическое окисление связано с отнятием от субстрата электронов и протонов. Роль же кислорода состоит в этом случае в регенерации окисленного состояния первичного акцептора водорода. По Баху, последовательность реакций в процессе могла бы выглядеть следующим образом:
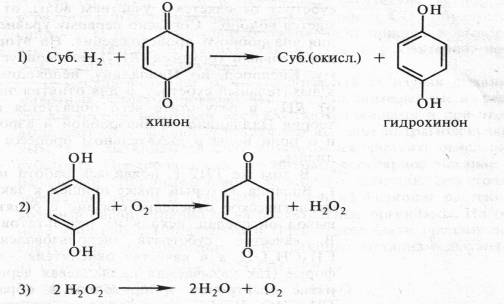
Эта вторая гипотеза Баха в дальнейшем была развита В. И. Палладиным в стройную теорию химизма дыхания. Занимаясь с 1903 по 1916 г. дыхательными пигментами (темнеющими на воздухе веществами тканей), Палладин нашел удачную искусственную модель, которая помогла расшифровать химизм дыхания. Это был известный краситель метиленовый синий. Если в пробирку с раствором метиленового синего поместить несколько прорастающих зародышей пшеницы и выкачать воздух (удалить кислород), то через некоторое время этот краситель полностью обесцветится. Если затем открыть пробирку и встряхнуть содержимое (т. е. дать кислород), то краситель снова синеет. Таким образом, при контакте с кислородом краситель окисляется, приобретая синий цвет, а живая ткань способна восстанавливать краситель, обёсцве чивая его. Причем окисление метиленового синего связано не с присоединением кислорода, а с отнятием водорода. Краситель восстанавливается за счет присоединения электронов и протонов.
На основании этих и других опытов Палладин в 1912 г. в статье «Значение дыхательных пигментов в окислительных процессах растений и животных» представил общую теорию химизма дыхания, разделив основное уравнение дыхания на анаэробную (1) и аэробную (2) части:

где R — это окрашенный дыхательный пигмент, способный отнимать водород от субстрата, a RH2 — бесцветный дыхательный хромоген. На первом, анаэробном, этапе дыхания глюкоза окисляется за счет отнятия водорода, который с помощью фермента (редуктазы) передается на дыхательный пигмент R (активация водорода!). По Палладину, дыхательный субстрат окисляется с участием воды, от которой тоже отнимается водород. Согласно первому уравнению весь С02 дыхания анаэробного происхождения. На втором, аэробном, этапе дыхательный хромоген (RН2) регенерирует в окисленную форму. Кислород, по Палладину, необходим не для включения в дыхательный субстрат, а для отнятия электронов и протонов от RН2, в результате чего образуется вода. В дальнейшем теория Палладина об анаэробной и аэробной фазах дыхания и о роли воды в дыхательном процессе полностью подтвердилась.
В том же 1912 г. появилась работа немецкого биохимика Г. Виланда, который также пришел к заключению о том, что биологическое окисление связано с отнятием водорода. Этот вывод он сделал, исходя из результатов модельных опытов. В качестве субстрата использовался этиловый спирт СН3СН2ОН, а в качестве окислителя — палладий в губчатой форме (так называемая палладиевая чернь). Происходило отнятие электронов и протонов и образовывался альдегид СН3СНО. Чтобы регенерировать восстановленный палладий и сделать реакцию долгоидущей, Виланд вводил хинон:
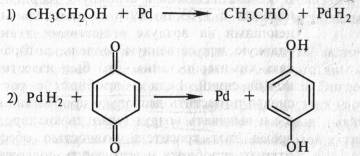
Эта реакция проводилась в небиологической системе и не предусматривала участие кислорода и воды.
Экспериментальные доказательства участия воды в окислении субстрата дыхания и роли 02 как конечного акцептора водорода были получены лишь в 1955 г. в работе Б. Б. Вартапетяна и A. Л. Курсанова. Опыты ставились с этиолированными проростками пшеницы, которым давали 180 в составе 02 или Н20, а через 2 ч анализировали содержание 180 в С02 дыхания
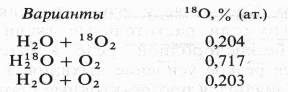
В состав С02 при дыхании тяжелый изотоп кислорода входил из меченой воды, но не из молекулярного кислорода.
Из всех приведенных выше данных следует, что окисление органических веществ в ходе дыхания связано с отнятием водорода и что теория Лавуазье о сходстве дыхания и горения не соответствует действительности.
3.Каталитические системы дыхания
Окисление дыхательных субстратов в ходе дыхания осуществляется с участием ферментов. Ферменты как белковые катализаторы, помимо свойств, присущих неорганическим катализаторам, обладают рядом особенностей: высокой актианостью, высокой специфичностью по отношению к субстратам и высокой лабильностью. Их пространственная организации зависящая от нее активность изменяются под действием внешних и внутренних факторов. Эти свойства обеспечивают возможность тонкой регуляции обмена веществ на уровне ферментов.
Типы окислительно-восстановительных реакций. Существуют четыре способа окисления, и все они связаны с отнятием электронов:
1) непосредственная отдача электронов, например:
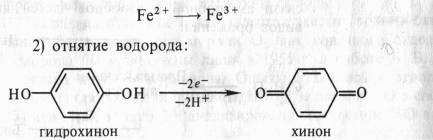
3) присоединение кислорода
![]()
4) образование промежуточного гидратированного соединения с последующим отнятием двух электронов и протонов:
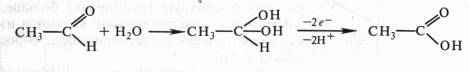
Оксидоредуктазы.
Поскольку окисление одного вещества (донора электронов и протонов) сопряжено с восстановлением другого соединения (их акцептора), ферменты, катализирующие эти реакции, называют оксидоредуктазами. Все они относятся к I классу ферментов:
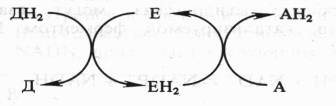
Донор (Д) отдает электроны и протоны, акцептор (А) принимает их, а энзим (Е) осуществляет реакцию переноса. Существуют три группы оксидоредуктаз: