Механизмы реализации путей гибели клетки
Т-, В, О - и NK-лимфоциты (lymphocytus). В крови взрослых людей они составляют 20-35% от общего числа лейкоцитов A,0-4,0109/л). Величина лимфоцитов в мазке крови значительно варьирует - от 4,5 до 10 мкм. Среди них различают малые лимфоциты (диаметром 4,5-6 мкм), средние (диаметром 7-10 мкм) и большие (диаметром 10 мкм и более). Большие лимфоциты встречаются в крови новорожденных и детей, у взрослых они отсутствуют. Для всех видов лимфоцитов характерно наличие интенсивно окрашенного ядра округлой или бобовидной формы, содержащего компактный гетерохроматин, и относительно узкого ободка базофильной цитоплазмы. В цитоплазме некоторых лимфоцитов содержится небольшое количество азурофильных гранул (лизосомы). Малые лимфоциты составляют большую часть (85-90 %) всех лимфоцитов крови человека. При электронной микроскопии в их ядрах выявляются небольшие впячивания; гетерохроматин расположен преимущественно по периферии ядра. В цитоплазме обнаруживаются везикулы, лизосомы, свободные рибосомы, полисомы, митохондрии, аппарат Гольджи, центриоли, небольшое количество элементов гранулярной эндоплазматической сети (рис.6).
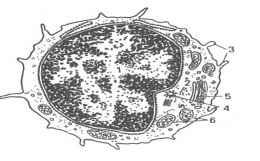
Рис.6. Ультрамикроскопическое строение лимфоцита (схема по Н.А. Юриной, Л.С. Румянцевой). I - ядро; 2 - рибосомы; 3 - микроворсинки; 4 - центриоль; 5 - аппарат Гольджи; 6 - митохондрии.
Среди малых лимфоцитов различают светлые и темные. Малые темные лимфоциты меньше светлых, имеют более плотное ядро, более узкий ободок базофильной цитоплазмы, обладающей высокой электронной плотностью. В цитоплазме расположено большое количество рибосом. Средние лимфоциты составляют около 10-12 % лимфоцитов крови человека. Ядра этих клеток округлые, иногда бобовидные с пальцевидным впячиванием ядерной оболочки. Хроматин более рыхлый, ядрышко хорошо выражено. В цитоплазме расположены удлиненные канальцы гранулярной эндоплазматической сети, элементы агранулярной сети, свободные рибосомы и полисомы, лизосомы. Центросома и аппарат Гольджи расположены рядом с областью инвагинации кариолеммы.
Кроме типичных лимфоцитов, в крови человека в небольшом количестве могут встречаться лимфоплазмоциты (около 1-2 %), которые отличаются концентрическим расположением вокруг ядра канальцев гранулярной эндоплазматической сети.
Основной функцией лимфоцитов является участие в иммунных реакциях. Однако популяция лимфоцитов гетерогенна по характеристике поверхностных рецепторов и роли в реакциях иммунитета.
Среди лимфоцитов различают три основных функциональных класса: В - лимфоциты, Т-лимфоциты и нулевые лимфоциты.
В-лимфоциты впервые были обнаружены в фабрициевой сумке птиц (bursa Fabricius), поэтому и получили соответствующее название. Они образуются у эмбриона человека из стволовых клеток - в печени и костном мозге, а у взрослого - в костном мозге. В-лимфоциты составляют около 30 % циркулирующих лимфоцитов. Их главная функция - участие в выработке антител, т.е. обеспечение гуморального иммунитета. Плазмолемма В - лимфоцитов содержит множество иммуноглобулиновых рецепторов. При действии антигенов В-лимфоциты способны к пролиферации и дифференцировке в плазмоциты - клетки, способные синтезировать и секретировать защитные белки - иммуноглобулины (Ig), которые поступают в кровь, обеспечивая гуморальный иммунитет.
Т-лимфоциты, или тимусзависимые лимфоциты, образуются из стволовых клеток костного мозга, а созревают в тимусе (вилочковая железа), что и обусловило их название. Они преобладают в популяции лимфоцитов, составляя около 70 % циркулирующих лимфоцитов. Для Т-клеток, в отличие от В-лимфоцитов, характерен низкий уровень поверхностных иммуноглобулиновых рецепторов в плазмолемме. Но Т-клетки имеют специфические рецепторы, способные распознавать и связывать антигены, участвовать в иммунных реакциях. Основными функциями Т-лимфоцитов являются обеспечение реакций клеточного иммунитета и регуляция гуморального иммунитета (стимуляция или подавление дифференцировки В-лимфоцитов). Т-лимфоциты способны к выработке лимфокинов, которые регулируют деятельность В-лимфоцитов и других клеток в иммунных реакциях. Среди Т-лимфоцитов выявлено несколько функциональных групп: Т-хелперы, Т-супрессоры, Т-киллеры. Т-хелперы (от англ. helper - помощник) - их главной функцией является усиление адаптивного иммунного ответа (рис.7). Активируют Т-киллеры, В-лимфоциты, моноциты, NK-клетки, презентируя им фрагменты чужеродного антигена при прямом контакте, а также гуморально, выделяя цитокины. Основным фенотипическим признаком Т-хелперов служит наличие на поверхности клетки молекулы CD4. Т-хелперы распознают антигены при взаимодействии их Т-клеточного рецептора (TCR) с антигеном, связанным с молекулами главного комплекса гистосовместимости 2 класса (MHC-II). Выделяют несколько подтипов Т-хелперов: Т-хелперы 0 (Th0) - "наивные" недифференцированные Т-хелперы; Т-хелперы 1 (Th1) - преимущественно способствуют развитию клеточного иммунного ответа, активируя Т-киллеры; основной выделяемый цитокин - интерферон-гамма; Т-хелперы 2 (Th2) - активируют В-лимфоциты, способствуя развитию гуморального иммунного ответа; продуцируют интерлейкины 4, 5 и 13; Т-хелперы 3 (Т-reg, Т-регуляторы, Т-супрессоры) - экспрессируют на поверхности молекулы CD25 и Foxp3, секретируют интерлейкин-10 и трансформирующий фактор роста-beta (TGF-beta) и супрессируют иммунный ответ: Т-хелперы 17 (Th17) - подтип Т-хелперов, который характеризуется продуцирует в больших количествах провоспалительный цитокин - IL-17. Показана роль Th17-клеток в развитии аутоиммуной патологии.
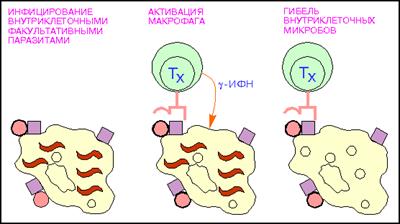
Рис.7: Активация макрофагов Т - хелпетами (кружки - поверхностный микробный антиген; красные квадраты - молекулы MHC класса II, волнистые линии - внутриклеточные паразиты.).
Регуляторные Т-лимфоциты, Т-супрессоры (англ. regulatory T cells, suppressor T cells, Treg) - центральные регуляторы иммунного ответа. Основная их функция - контролировать силу и продолжительность иммунного ответа через регуляцию функции Т-эффекторных клеткок (Т-хелперов и Т-цитотоксических клеток). Эти клетки экспрессируют FOXP3 - транскрипционный фактор, регулирующий транскрипцию генов, ответственных за дифференцировку Т-клеток и экспрессию цитокинов и других факторов, участвующих в супрессии иммунного ответа. Часто эти клетки так и обозначают, как FOXP3+ регуляторные Т-клетки (FOXP3+ Treg cells). Кроме того, важным маркером Т-регуляторных клеток является экспрессия на их поверхности рецептора к цитокину IL-2 - CD25, соответственно это обозначают как CD25+ клетки. Помимо этих основных маркёров Treg клетки на своей мембране экспрессируют CD62L, различные изоформы мембрано-связанной фосфатазы CD45. Различают несколько разных типов регуляторных Т-клеток: естественные Т-регуляторные клетки (T-reg1) и индуцибельные Т-регуляторные клетки (iT-reg). Индуцибельные Т-регуляторные клетки образуются под влиянием различных факторов на периферии, например, в региональных лимфатических узлах. Для супрессии иммуного ответа Тreg клетки выделяют цитокины: TGF-beta, IL-10, IFNgamma, IL-35, а также экспрессируют на своей поверхности рецептор CTLA-4. Существуют несколько механизмов супрессии: прямой, при непосредственном контакте между клетками, и дистантный, осуществляющийся на расстоянии - например, через растворимые цитокины. Мишенями действия Тreg клеток являются как Т-эффекторные клетки, так и дендритные клетки, ответственные за презентацию антигена и активацию Т-клеток. При дистантном механизме цитокины, выделяемые Тreg клетками, например, трансформирующий ростовой фактор бета (TGF-beta), связываются со своими рецепторами на поверхности Т-эффекторных клеток и ингибирует их активацию, тем самым супрессируя иммунный ответ. Важным механизмом также служит захват IL-2 при помощи CD25 - рецептора к IL-2 и секвестрация рецептора у эффекторных Т-клеток, что препятствует активации после связывания комплекса MHC с антигеном, так как известно, что IL-2 является основным аутокринным стимулирующим фактором, поддерживающим диффернцировку и клональную экспансию Т-клеток. Тreg клетки, взаимодействуя с рецептором CD86 на дендритных клетках при помощи CTLA-4, способны ингибировать функцию активации дендритными клетками Т-клеток. При прямом механизме супрессии Treg взаимодействуют с эффекторными Т-клетками и гранзим B действует через перфорины, образующие канал, вызывая апоптоз в этих клетках, тем самым элиминируя активные Т-клетки. Цитотоксические Т-лимфоциты непосредственно контактируют с чужеродными клетками и разрушают их. Клеточный иммунитет опосредован цитотоксическими Т-лимфоцитами и Т-хелперами. Многие микроорганизмы живут внутри клеток в недосягаемости для гуморальных антител. Чтобы справиться с внутриклеточными паразитами, возникла обособленная система приобретенного имунитета, основанная на функционировании отдельной субпопуляции лимфоцитов, а именно, цитотоксических T-лимфоцитов (T-киллеров). Эти клетки отличаются очень широкой специфичностью. Поскольку T-лимфоциты должны взаимодействовать с содержащими паразитов клетками, они могут узнать антиген только в том случае, если он экспрессирован на поверхности инфицированной клетки. Как и B-лимфоциты, каждая T-клетка имеет специфический рецептор, который распознает этот антиген. (Рецептор T-лимфоцита структурно отличается от молекулы мембраного иммуноглобулина - рецептора B-лимфоцитов). При этом T-лимфоциты узнают антиген на поверхности клетки в комплексе с клеточным маркером: молекулами MHC класса I. В процессе распознавания поверхностного антигена цитотоксический T-лимфоцит вступает в теснейший контакт со своей мишенью и уничтожает ее до начала репликации. Кроме того, он продуцирует гамма-интерферон, который ограничивает проникновение вируса в соседние клетки, особенно в тех случаях, когда вирус является слабым индуктором альфа-интерферона и бета-интерферона Цитотоксические T-лимфоциты относятся к субпопуляции T8 (или Ly2) и представляют популяцию T-эффекторов и играют важнейшую роль в уничтожении клеток, зараженных вирусами, бактериями, а возможно, участвуют и в предполагаемом надзоре за появлением опухолевых клеток. Большинство цитотоксических T-клеток (Tц) относится к субпопуляции CD8+ и распознает антиген, презентированный в ассоциации с молекулами MHC класса I, но меньшая их часть (примерно 10%), относящаяся к субпопуляции CD4+, способна распознавать антиген в ассоциации с молекулами MHC класса II. Для активации и поражения мишени Tц обладают несколькими механизмами воздействия. Один из них - это передача сигналов при непосредственном клеточном контакте через поверхностные структуры, другой - непрямая сигнализация с помощью цитокинов. Кроме того в цитоплазме многих цитотоксических клеток обнаружены гранулы с белками, высвобождение которых вблизи цитоплазматической мембраны клетки-мишени вызывает ее повреждение. Цитотоксические T-лимфоциты развиваются из предшественников. Предшественники цитотоксических клеток активируются комплексом антигена и молекул MHC класса I, размножаются и созревают под действием IL-2, а также еще плохо идентифицированных факторов дифференцировки, продуцируемых T-хелперами.