Молекулярные основы наследственности
Клеточный цикл
События, происходящие в период от одного клеточного деления до другого, называются клеточным циклом. Фаза митоза цикла охватывает период деления и хромосом, и клеток. После расхождения клеток каждая дочерняя клетка вступает в период повышенной биосинтетической активности - в так называемую Gj-фазу. Gj-фаза заканчивается перед началом удвоения хромосом, или, в молекулярных терминах, с началом дупликации хромосомной ДНК; период репликации генома называется фазой синтеза. С момента завершения S-фазы в клетках инициируются события, характерные для митотической профазы,-части цикла, называемой Gj-фазой. В конце концов опять начинаются митоз и цитокинез, и цикл повторяется. Как правило, Gr, S - и G2-периоды, вместе составляющие интерфазу, занимают около 90% времени клеточного цикла, а М-фаза - менее 10%. Полное время прохождения клеточного цикла в клетках разного типа сильно варьирует в зависимости от условий роста. Основным показателем продолжительности всего цикла является продолжительность Gj-фазы. Например, покоя:
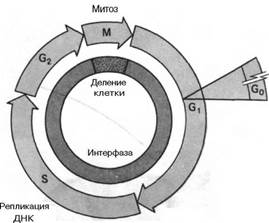
Клеточный цикл: митоз и цитокинез составляют М-фазу цикла, кульминацией которой является образование двух дочерних клеток. Каждая дочерняя клетка вступает в G1-период интерфазы и может начать новый клеточный цикл. За периодом G1 следует S-фаза, во время которой ДНК и хромосомы дуплицируются, и далее - фаза G2. Начало митоза означает конец интерфазы. Покоящиеся клетки задерживаются в фазе G1 и, как говорят, находятся в фазе G0. Обычно эукариотические клетки, которые не остановились в фазе G0, завершают цикл за 24 ч.
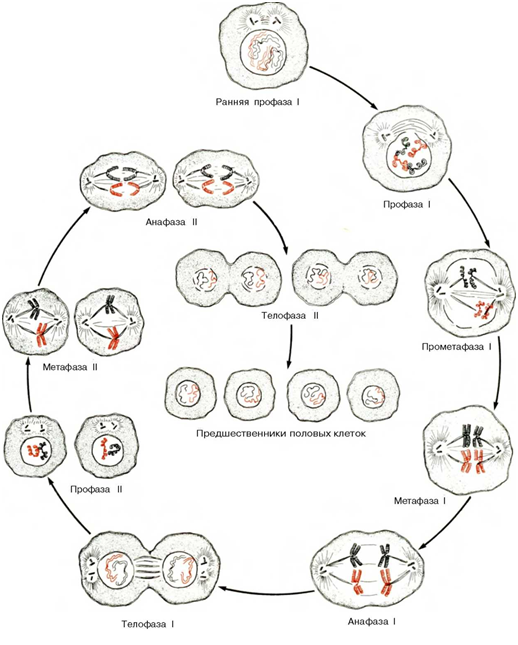
Мейоз: этапы деления диплоидной клетки на четыре гаплоидные дочерние клетки. Этот процесс отличается от митоза тем, что включает два клеточных деления и только один "раунд" репликации хромосом. На схеме показаны две пары гомологичных хромосом. Во время интерфазы хромосомы имеют вид тонких диффузных нитей. После репликации сестринские хроматиды остаются тесно связанными и начинают конденсироваться, что указывает на начало профазы. Затем гомологичные пары сестринских хро-матид приходят в тесное соприкосновение, образуя тетрады; этот процесс называется синапсисом. Начало мейотической метафазы I характеризуется дальнейшей конденсацией хромосом и дезинтеграцией ядерной мембраны. В анафазе I члены гомологичной пары сестринских хроматид начинают перемещаться к разным полюсам удлиняющейся клетки. К концу телофазы I и клеточного деления I образуются две дочерние клетки, в каждой из которых имеется по одной гомологичной паре сестринских хроматид. Второй раунд клеточного деления происходит без дополнительной дупликации хромосом и начинается с профазы II, с переходом в метафазу II. В стадии анафазы II две сестринские хроматиды, которые оставались до этого момента вместе, начинают перемещаться к противоположным концам удлиняющейся клетки. После телофазы II и клеточного деления II образуются четыре гаплоидные клетки - предшественники половых клеток. В каждую дочернюю клетку попадает только по одной хромосоме из исходных гомологичных пар.
 |
Образование яйцеклеток и сперматозоидов подразумевает уменьшение нормального числа хромосом ровно вполовину; этот процесс называется мейозом. Гаметы, или половые клетки, гаплоидны, т.е. в них содержится по одному члену каждой пары гомологичных хромосом, и, таким образом, только половинное число хромосом каждого из родителей попадает во все другие, соматические, клетки организма потомка. Распределение хромосом в мейозе происходит случайно, поэтому любой из членов гомологичной пары может оказаться во вновь образовавшихся зародышевых клетках.
При оплодотворении гаплоидные наборы хромосом сперматозоидов и яйцеклеток объединяются. Таким образом восстанавливается полный набор гомологичных хромосомных пар, каждый из членов которых произошел из яйцеклетки и из сперматозоида соответствующих родителей. Диплоидное состояние оплодотворенной яйцеклетки поддерживается далее во всех соматических клетках механизмом митотического деления. Иногда зрелые организмы могут развиться из неоплодотворенных гаплоидных яйцеклеток или из оплодотворенных яйцеклеток с неполным набором родительских хромосом. Как уже отмечалось, любой из членов гомологичной пары может попасть в функциональную гамету. В зрелую яйцеклетку или сперматозоид попадает по одному члену каждой пары в процессе редукции числа хромосом в мейозе.
Строение хромосом
Легче всего наблюдать метафазные хромосомы. Под микроскопом их фотографируют или зарисовывают. В этой стадии хромосомы наиболее сконденсированны и образуют дискретные структуры. У многих организмов индивидуальные хромосомы и их гомологи легко различимы по размеру и форме. Каждая метафазная хромосома действительно состоит из двух идентичных частей, называемых сестринскими хроматидами, поскольку дупликация хромосомной ДНК протекает как раз перед метафазой, в S-фазе клеточного цикла.
У хромосомы имеется перетяжка, называемая центромерой. Положение центромеры для каждой хромосомы строго определено. С центромерой связаны специфические хромосомные функции; это последняя точка, соединяющая плечи сестринских хроматид перед полным расхождением при митотическом или II мейотическом делении. Сами плечи имеют вид
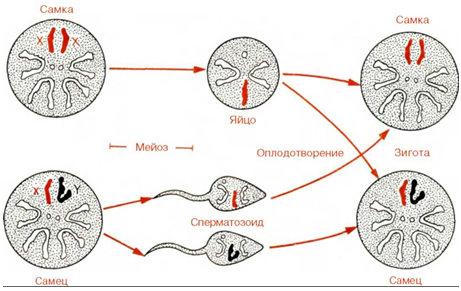 |
Образование гаплоидных гамет при мейозе и слияние двух гамет с образованием диплоидной клетки при оплодотворении. Обратите внимание на то, что у D. melanogaster, рассмотренной здесь в качестве примера, как и у других организмов, включая млекопитающих, две половые хромосомы у самца не гомологичны друг другу. При мейозе формируются два типа сперматозоидов, из которых один несет Х-, а другой - Y-хромосому. У самок, несущих пару Х-хромосом, в результате мейоза образуются гаметы одного типа. Пол потомков зависит от того, какую из хромосом - X или Y - несут оплодотворяющие сперматозоиды. У некоторых организмов негомологичную, определяющую пол хромосому несет самка.
Различие между областью центромеры и плечами хромосом становится очевидным после обработки определенными красителями. После окрашивания центромеры выглядят более плотными и компактными по сравнению с плечами. Такие плотные, интенсивно окрашиваемые хромосомные области называются гетерохроматиновыми. Гетерохроматин центромеры можно наблюдать после окрашивания даже в плохо различимых интерфазных хромосомах. Другие, негетерохроматиновые области хромосом принято называть эухроматиновыми. Эухроматиновые области окрашиваются гораздо менее интенсивно, чем гетерохроматиновые.