Морфология колонии у гидроида obelia loveni
Показатели структуры колоний Obelia loveni (Allm.) (значения символов указаны в тексте)
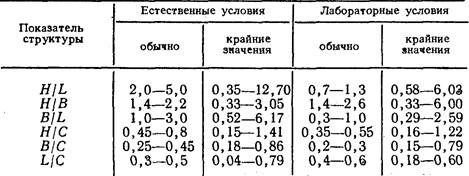
В наших опытах показатели структуры были рассчитаны на основании 86 картирований 19 колоний, культивируемых в естественных условиях, и 55 картирований 9 колоний, содержащихся в условиях лаборатории в течение двух месяцев — июля и августа. Результаты показали, что колонии в естественных и лабораторных условиях имели разное строение (таблица). Сравнение морфологии колоний двух серий показало, что они различаются степенью разветвленности побегов, их максимальной высотой, расстояниями между побегами и степенью развития гидроризы. Старые побеги в естественных условиях были более высокими и разветвленными, чем в лабораторных. Однако гидрориза была длиннее у лабораторных колоний, и расстояния между побегами здесь были достоверно больше.
В течение двух месяцев показатели структуры менялись без видимой закономерности, не выявляя месячной ритмики, обнаруженной ранее у Dynamena pumila (Марфенин, 1977).
Анализ морфологических показателей. Подробное изучение морфологии колонии дает возможность существенно расширить список морфологических показателей, используемых в систематике семейства Campanulariidae (Cornelius, 1982). В то же время в ряде случаев представляется возможным объяснить их функциональную роль в жизнедеятельности колонии. Это позволяет несколько приблизиться к оценке сравнительной значимости морфологических показателей, столь важной при моделировании формы любого организма. Мы полагаем, что колониальные гидроиды особенно перспективны как объекты моделирования из-за относительной простоты количественного описания их формы. Первая попытка подобного моделирования была предпринята сравнительно давно (Braverman. Schrandt, 1967) и выявила недостаток числа количественных показателей, используемых в описании колонии у гидроидов.
Мы надеемся, что приведенные ниже показатели окажутся полезными как в систематике гидроидов, так и при моделировании формы побега и колонии в целом.
1. Диаметр ствола побегов, ветвей, ножек гидрантов и столонов — основополагающая характеристика колонии, особенно в семействе Campanulariidae, так как вся колония фактически “построена из трубок”. Диаметр перисарка косвенно указывает на эффективность использования клеточного материала (чем больше диаметр, тем больше живых тканей в единице колонии), а также тесно связан с жесткостью конструкции. У О. loveni относительно тонкие стволы и гидрориза. Такие колонии гибки, но не обладают необходимой жесткостью для того, чтобы противостоять течению воды.
2. Переплетение ветвей побегов способствует усилению их общей жесткости. Переплетение достигается за счет значительного угла отхождения боковых ветвей от ствола и других ветвей, а также за счет интенсивного ветвления.
3. Прямолинейный рост гидроризы и ее ветвление под прямым углом способствуют относительно быстрому распространению по субстрату и освоению его.
4. Локализация мест ветвления гидроризы вблизи оснований побегов приводит к увеличению прочности прикрепления колоний к субстрату, что особенно важно при увеличении размеров и соответственно парусности побегов. Специальных расширений столонов под побегами, свойственных, например, семейству Sertulariidae, у Obelia не выработалось. Такие расширения столонов связаны с закреплением мест ветвления гидроризы исключительно под побегами.
5. Отсутствие анастомозов в гидроризе тесно скоррелировано с умеренностью ее ветвления, а это означает преобладание стратегии прямолинейного роста столонов и захвата новых площадей субстрата перед стратегией сдерживания экстенсивного роста и наиболее полным использованием занятой территории. Последнее присуще таким гидроидам, как Clava и Hydractinia, обитание которых обычно приурочено, по нашим наблюдениям, к определенным местам субстрата.
6. Доля междоузлиq и гидрантов, приходящихся на боковые ветви побега, демонстрирует значение ветвления, которое способствует концентрации гидрантов, т. е. увеличению их числа на единицу площади, занимаемой колонией. Доля гидрантов в стволе и боковых ветвях разного порядка показывает роль разветвления колонии и характер распределения функционирующих гидрантов в кроне, которое может быть равномерным или, к примеру, в основном периферическим — по поверхности кроны.
7. Высота побега, ширина кроны, длина междоузлия, число порядков разветвления, угол отхождения боковых ветвей от ствола, угол наклона боковых ветвей к плоскости “гидрант — ствол”, частота ветвления, продолжительность роста боковых ветвей — основные количественные показатели, с помощью которых можно точно моделировать форму побега.
8. Расстояния между побегами, частота ветвления гидроризы, место и угол отхождения боковых столонов от материнского — показатели, необходимые для моделирования колонии в целом.
9. Продолжительность жизненного цикла гидранта и число возможных жизненных циклов на одном междоузлии — функциональные показатели, необходимые для моделирования колонии в развитии.
Анализ тотальной морфологии колонии. В систематике гидроидов до сих пор используются почти исключительно показатели морфологии отдельных “типичных” фрагментов, как правило зооидов. Морфология побега описывается весьма приблизительно почти без использования количественных показателей. Естественно, что при таком подходе различия между побегами не заметны, а колония выглядит как простая сумма “одинаковых” побегов с одинаковыми зооидами.
Более подробное изучение целых колоний с применением методики их количественного картирования позволяет выявить различия между побегами и обнаружить закономерности в строении колонии. Различия между побегами выражаются в числе междоузлии и числе гидрантов, в разветвленности, а следовательно, и в числе верхушек роста, в наличии и числе гонозооидов.
Строение побегов меняется в пределах ветви гидроризы, на которой они находятся, постепенно и закономерно от одного ее конца к противоположному, что создает эффект морфологической полярности, или градиента, впервые отмеченный А. Г. Карлсеном (1982) на Dynamena pumila.
Ветви гидроризы с побегами у О. loveni соединены между собой определенным образом, так, что общее строение колонии можно назвать “центробежным”. Морфологическая полярность ветвей гидроризы вследствие “центробежного” строения колонии отражается и в ее общем строении. В колонии можно различить три зоны: центральную (или зону депрессии) и расположенные концентрически вокруг нее среднюю зону (или зону воспроизводства) и периферическую зону (роста).Часто зона депрессии еще отсутствует, и тогда можно выделить только две зоны — периферическую и зону воспроизводства, которая в этом случае занимает центральное положение. Подобное подразделение колонии на две зоны было предложено нами ранее для Dynamena pumila (Марфенин, 1973). Теперь мы убедились, что это лишь частный вариант, а более общий — дифференциация колонии на описанные выше три морфофункциональные зоны.