Муха
У D. melanogaster найдено шесть факторов фертильности самцов (kl-5, kl-3, kl-2, kl-1, ks-I и ks-2 нарис. З) Из них три очень больших: kl-5, kl-3 и ks-1 — занимают по 10% Y-хромосом каждый, то есть примерно по 4000 т.п.н.
Интересно проявляется активность факторов фертильности у дрозофилы. В 1961 году три немецких ученых (G.F. Меуег, О. Hess, W. Beermann) описали особые нитевидные структуры в ядрах развивающихся сперма тоцитов D. melanogaster, которые впоследствии стал называть петлями (рис. 5). Такие структуры нашли фактически у всех 50 изучаемых видов дрозофилы. Показано, что петли - это декомпактизованные, а следовательно, активные участки Y-хромосом. В них синтезируется РНК и накапливаются белки. Каждая петля ядре данного вида дрозофилы имеет характерные размеры, ультраструктуру и внешний вид (см. рис. 5). У других видов морфология набора петель другая.
О том, что петли формируются из материала Y-xpомосомы, свидетельствуют следующие факты.
1. У самцов, не имеющих Y-хромосомы (ХО), нет и петель, а у особей с двумя Y-хромосомами (XYY)ониприсутствуют в двойном наборе. Если происходит делеция части Y-хромосомы, обнаруживаются не все петли. В линиях с дупликациями частей Y-хромосом число петель соответственно увеличивается.
2. У межвидовых гибридов морфология петель такая же, как и у вида - донора Y-хромосомы.
Более детальный анализ показал, что гены ферментильности самцов локализованы в петлях.
1.Сначала были установлены корреляциимежду числом генов и петель. Затем, используя хромосомные – перестройки, установили прямое соответствие в их локализации. Так, фактор kl-5 соответствует петле А, поскольку и петля, и фактор располагаются между точками разрывов одних и тех же перестроек (см. рис. 3). Фактор kl- 3 расположен в петлеB, ks-1 - в петле С.
2. При удалении делециями хотя бы одной петли самец становится стерильным.
После получения клонов ДНК из Y-хромосом дрозофил появилась возможность анализа молекулярной организации этой хромосомы. Общая длина петель составляет около 1000 мкм, или 1/12 всей длины ДНК в Y-хромосоме. Функции остальных 11/12 пока неизвестны. В состав ДНК Y-хромосомы входят два типа повторенных последовательностей.
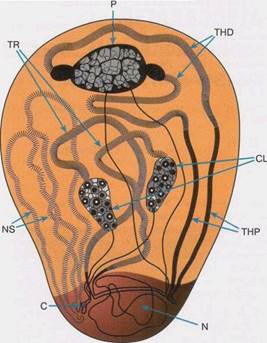
Рис. 3. Общий вид ядра спермотоцита у самца Drosofiliahydei(из [1], с.62). TR, P, THD, CL, THP, NS – названия петель, С – центромера, N - ядрышко
Вывод:
Таким образом, муха Дрозофила играет большую роль, как объект генетических исследований. Исследование ее генов принесло известность многим генетикам. Изучая гены Дрозофилы, ученые открыли много законов, таких как: закон Моргана - сцепленного наследования генов и закон Менделя. К тому же у Дрозофилы нашли много различных генов, отвечающих за наследование некоторых признаков. В последствии многие из этих открытий применялись к людям. И если бы не было этих мух, ученые еще долго бы не открыли законы наследования у людей.
Таблица 1.
|
Свойства |
Эухроматин |
Гетерохроматин |
|
Доля генома |
67% |
33% |
|
Расположение в хромосомах |
Плечи хромосом |
В прицентромерных областях, вся Y-хромосом |
|
Состояние компактности в клеточном |
В ходе митотического и мейоти- |
На протяжении всего клеточного цикла |
|
Цикле |
ческого делений | |
|
Компактизующее влияние на приближен |
Не оказывает |
Участки эухроматина, приближенные к гетеро |
|
ные участки хромосом (эффект положе |
хроматину, также становятся компактными, | |
|
ния мозаичного типа) |
гены в них инактивируются | |
|
Способность объединяться с другими |
Не отмечена |
Гетерохроматиновые участки объединяйте; |
|
районами хромосом |
образуя хромоцентры | |
|
Образование хромосомных перестроек |
Обычная частота обнаружения |
Повышенная частота обнаружения |
|
Расположение в клеточном ядре |
По всему объему ядра |
Главным образом на ядерной оболочке |
|
Время синтеза ДНК в клеточном цикле |
Первые 3/4 периода синтеза ДНК |
Последняя половина S-периода. Завершени |
|
в интерфазе (S-периода) |
процесса репликации ДНК сильно задержано | |
|
Дифференциальная окраска специфичес |
Отсутствие окраски |
Интенсивная окраска |
|
кими красителями (С-окраска) | ||
|
Фракции ДНК по степени повторенности |
-90% уникальных последователь |
Основная масса ДНК представлена высокопое |
|
ностей и -10% умеренно повто |
торенными фракциями, в меньшей степен | |
|
ренных |
умеренными повторами и совсем мало уникал! | |
|
ных последовательностей | ||
|
Наличие особых компактизующих бел |
Почти отсутствует |
Обильно присутствует по всему гетерохромг |
|
ков, например белка НР1 |
тину | |
|
Варьирование количества материала в |
Заметное варьирование не обна |
Варьирование количества гетерохроматина un |
|
хромосомах |
ружено |
роко представлено в каждой хромосоме |
|
Генетическое содержание |
Основная часть всех генов генома |
Гены почти отсутствуют |
|
локализована в эухроматине |