Обнаружение единичных нуклеотидных замен в ДНК расщепление РНКазой и денатурирующий градиентный гель-электрофорез
1.2.2 Практическое использование ДГГЭ
Приведенные здесь схемы ДГГЭ обеспечивают более высокую разрешающую способность и эффективность обнаружения мутаций за счет использования гетеродуплексов. Их основные этапы приведены на рис. 4. Исследуемые препараты ДНК отжигают с меченым одноцепочечным ДНК-зондом, и если в них произошла замена нуклеотида, образовавшийся гетеродуплекс будет иметь однонуклеотидный неспаренный участок. Далее проводят электрофорез в денатурирующем геле с последующей радиоавтографией, используя в качестве контроля гомодуплекс ДНК дикого типа. Таким образом, в этом случае не требуется стадии блотинга геля. Кроме одноцепочечных ДНК-зондов можно использовать асимметрично меченные двухцепочечные ДНК-зонды, а также меченые одноцепочечные РНК-зонды для тестирования ДНК в гибридах РНК–ДНК или РНК в РНК–РНК-гибридах.
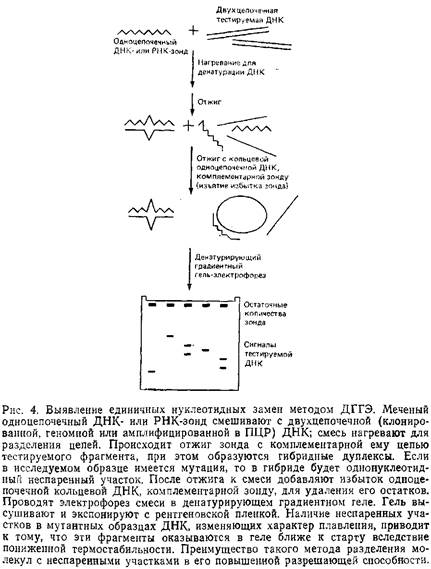
С помощью ДГГЭ, также как и при РНКазном расщеплении, можно исследовать фрагменты нуклеиновых кислот длиной от 100 до 1000 п.н. Верхний предел значений длин в некоторой степени определяется необходимостью использовать полиакриламидный гель. Подвижность в нем фрагментов ДНК длиной более 1000 п. н. резко снижается, а это значительно увеличивает время, необходимое для их электрофоретического разделения. Еще более серьезную проблему представляет собой характер плавления длинных фрагментов. Известно, что чем длиннее фрагмент, тем. больше в нем доменов плавления. При прохождении такого фрагмента через гель очень быстро наступает резкое снижение его подвижности, обусловленное плавлением одновременно большого числа доменов, что делает доступной для обнаружения замены лишь незначительную часть длинного фрагмента. В силу вышеуказанных причин мы стараемся работать с фрагментами, длина которых не превышает 1000 п. и. Поскольку для большинства фрагментов ДНК более половины их длины приходится на легкоплавкие домены, то денатурирующий электрофорез фрагмента в 1000 п. и. позволяет тестировать на нуклеотидные замены примерно 500 нуклеотидов. Для повышения информативности анализа можно использовать ДНК-зонд длиной 1000–2000 п.н.; отжигать его с геномной ДНК, обрабатывать соответствующими рестриктазами и получать при этом два-три фрагмента оптимального размера.
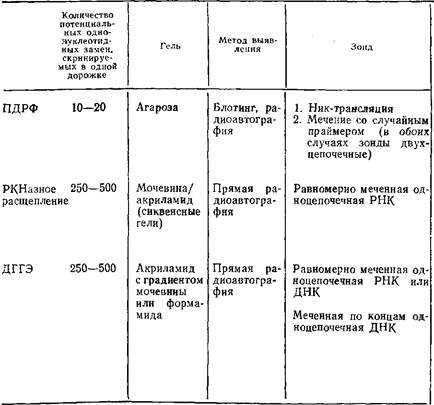
Таблица 1. Сравнительная характеристика методов градиентного анализа ПДРФ, рестриктазного расщепления и денатурирующего гель-электрофореза

1.3. Полимеразная цепная реакция
РНКазное расщепление и ДГГЭ можно использовать для непосредственного исследования фрагментов геномной ДНК, минуя стадию клонирования. Работая с равномерно меченными зондами, обладающими высокой удельной активностью, можно любым из этих методов получить желаемый результат, имея изначально 5–10 мкг геномной ДНК человека и проводя радиоавтографию 24 ч. Чем проще организм, тем выше чувствительность методов. Хотя получаемые результаты в большинстве своем можно оценить высоко, непосредственное использование суммарной геномной ДНК ставит перед исследователем ряд проблем. Это, во-первых, нередко низкое отношение сигнала к фону, особенно при РНКазном расщеплении. Во-вторых, активность равномерно меченных фосфором зондов должна быть настолько высокой, чтобы использовать их в течение дня или двух. В-третьих, при проведении некоторых анализов, особенно в случае множественных тестов, лимитирующим фактором может стать количество геномной ДНК. Любой тест тем предпочтительнее, чем меньшее количество ДНК требует.
В 1985 г. был описан принципиально новый метод, позволяющий в миллион раз амплифицировать интересующие последовательности в препарате геномной ДНК, – полимеразная цепная реакция, ПЦР. Идея метода и ее воплощение очень просты. Сначала синтезируются два дезоксиолигонуклеотида длиной 20–30 оснований, представляющие собой концевые последовательности интересующего фрагмента ДНК. Полярность выбрана так, чтобы после отжига их направления 5'-3' были обращены друг к другу. Избыточные количества этих олигонуклеотидов смешивают с геномной ДНК, и смесь нагревают для денатурации последней. Снижение температуры приводит к реассоциации олигонуклеотидов с гомологичными участками геномной ДНК. Затем проводят наращивание цепи при участии ДНК-полимер азы и дезоксирибонуклеотидтрифосфатов. Такая последовательность реакций денатурации, реассоциации и наращивания цепи повторяется 20–30 раз. Уже после двух циклов среди продуктов реакции появляются фрагменты ДНК, точно совпадающие по длине с исходным фрагментом, ограниченным олигонуклеотидами. Эти фрагменты служат матрицей для последующих реакций и идентичны большинству конечных продуктов. Процесс является по существу цепным, так как продукты данной реакции служат матрицей для последующих реакций. Количество вновь образующейся ДНК возрастает в геометрической прогрессии, поэтому за 20 циклов при 100% – ной эффективности каждого из них можно получить 220 молекул. На практике эффективность каждого цикла амплификации составляет 20–50%, т.е. при проведении достаточного числа циклов можно добиться увеличения количества специфической последовательности кратного миллиону.
Если при амплификации геномной ДНК позвоночных в качестве праймеров для ПЦР используют олигонуклеотиды длиной 20 нуклеотидов и более, то процесс этот довольно специфичен и амплифицируется только один фрагмент ДНК – Однако иногда среди продуктов реакции наблюдается накопление фрагментов, происхождение которых трудно объяснить. А поскольку эти фрагменты могут мешать последующему анализу, то рекомендуется провести еще одну серию амплификации, с использованием другого набора олигонуклеотидных праймеров. Этот метод, именуемый «nested oligo», состоит в следующем: продукт первичной полимеразной цепной реакции используется в качестве матрицы в последующих раундах ПЦР, но уже с двумя другими олигонуклеотидами, имеющими гомологии с участками ДНК внутри первичного амплифицированного фрагмента. Такая процедура позволяет получить большое количество индивидуальной последовательности ДНК, несколько более короткой, чем исходный фрагмент, и использовать ее для последующего анализа.
Имея исходно менее 1 мкг суммарной геномной ДНК позвоночных, в ПЦР можно получить несколько микрограмм специфического фрагмента. Хорошо амплифицируются фрагменты до 2000 п. н. В одной реакции можно амплифицировать одновременно и несколько фрагментов. Амплификация специфических фрагментов с помощью ПЦР может применяться и в диагностике, и при клонировании. Левинсон и Гитшейер использовали амплифицированную в ПЦР геномную РНК и РНКазное расщепление для выявления однонуклеотидных замен в гене фактора VIII, обусловливающих Х-сцепленную гемофилию А человека. Мы объясняем, как применять ПЦР в сочетании с РНКазным расщеплением и с ДГГЭ для обнаружения однонуклеотидных замен в препаратах геномной ДНК.
