Определение таксономического состава и вертикального распределения организмов дрифта беспозвоночных
Большинство организмов сносятся в поверхностном горизонте воды (табл. 2), что еще раз доказывает тот факт, что дрифт не является полностью пассивным процессом (Леванидов, Леванидова, 1981).
Более высокая плотность животных в верхних слоях воды объясняется их целенаправленными миграциями, которые могут быть спровоцированы столкновениями конкурирующих особей. Это может быть связано с вылетом ряда групп амфибиотических насекомых, которому предшествует активный предимагинальный дрифт (Леванидов, Леванидова, 1981).
Пассивное всплытие организмов происходит вместе с фрагментами перифитонных матов (сем. Naididae) или увлечением их с турбулентными токами воды (личинки и взрослые жуки Elmidae) (Паньков, 2007).
Литературные данные о вертикальном распределении организмов дрифта довольно противоречивы. Так в мощном предгорном потоке (р. Хор) плотность всплывающих организмов в верхнем горизонте была в 4 – 5 раз выше, чем в придонном (Леванидов, Леванидова, 1981).
В небольшой горной реке Щугор, напротив, наибольшая численность дрейфующих организмов приурочена к придонному горизонту (Шубина, 1986). Мигрирующие беспозвоночные преобладают в придонном горизонте и в р. Волге (Аристовская, 1945).
Таким образом, вертикальное распределение организмов дрифта еще не достаточно изучено и нуждается в дальнейшем исследовании.
4.3 Суточная динамика интенсивности дрифта
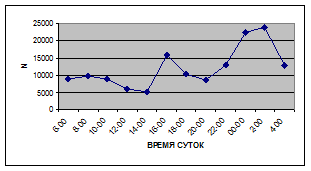
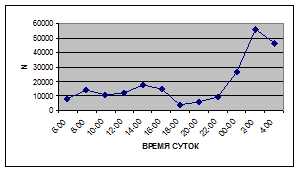
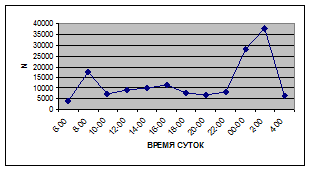
Суточная динамика интенсивности дрифта донных беспозвоночных р. Сылвы описывается кривой довольно сложной конфигурации, на которой могут быть выделены максимумы и минимумы первого и второго порядков (рис. 3).
Рассмотренная кривая на уровне элементов первого порядка имеет одновершинную форму с наивысшими показателями с двенадцати до двух часов ночи и низкими показателями в дневные часы.
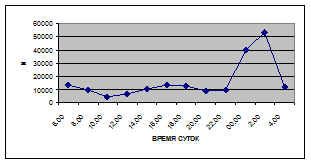
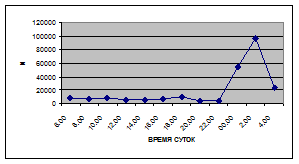
а) 29 июня 2005 г. д) 19 июня 2006 г.
б) 06 июля 2005 г. е) 11 июля 2006 г.
в) 16 июля 2005 г. ж) 18 июля 2006 г.
г) 18 июля 2005 г. з) 27 июля 2006 г.
Рис. 3. Суточная динамика интенсивности дрифта (N, экз/ м2 в час, водная фракция)
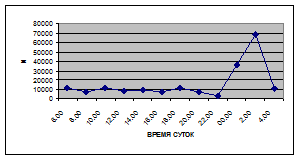
Важной характеристикой дрифта является амплитуда колебаний его параметров в течение суток. В р. Сылве максимальная интенсивность дрифта (Nmax) превосходит минимальную (Nmin) в 4,5 – 23,4 раз.
Минимальные показатели интенсивности дрифта приурочены к дневному времени суток, в основном, с четырех часов утра до восьми часов вечера. Они изменяются в пределах от 7,2 до 21,4 тыс. экз/ м² в час.
В этот период мигрируют животные с разной склонностью к дрифту. Это могут быть и животные, сравнительно легко расстающиеся с бенталью и животные, в целом не расположенные к вертикальным миграциям.
К ним относятся водные клещи Hydracarina и клопы Micronecta sp., ракообразные Corophium curvispinum Sars, 1895 и Ostracoda sp., личинки амфибиотических насекомых Trichoptera, Ephemeroptera, Diptera и другие (прилож. 1, 5, 9, 13, 17, 21, 25, 29).
Далее, с десяти до двенадцати часов ночи, наблюдается увеличение интенсивности дрифта до 40,5 тыс. экз/ м² в час. Это происходит за счет увеличения миграционной активности амфибиотических насекомых: личинок и куколок Сhironomidae, Leptoceridae, Baetidae (Cloeon luteolum (Muller, 1776)).
К двум часам ночи интенсивность дрифта возрастает до максимальных значений в суточном цикле, в среднем до 63,8 тыс. экз/ м² в час. Эти показатели обусловлены подъемом в толщу воды массовых групп животных. К ним относятся личинки и куколки Сhironomidae, Trichoptera, Cloeon luteolum, Baetis scambus и другие таксоны (прилож. 1, 5, 9, 13, 17, 21, 25, 29).
Приуроченность наивысших показателей интенсивности дрифта к первой половине ночи отмечена и в литературе (Леванидов, Леванидова, 1981; Богатов, 1983; Шубина, 1986; Задорина, 1987б).
Именно в эти часы дрифт достигает не только максимальной интенсивности, но и максимального разнообразия – в пробах отмечены все зарегистрированные таксономические группы (прилож. 1, 5, 9, 13, 17, 21, 25, 29).
Пик интенсивности дрифта обусловлен возрастанием миграционной активности всех участвующих в дрифте групп. Наибольшая интенсивность дрифта в эти часы зарегистрирована у личинок Ephemeroptera – 20,1 тыс. экз/ м², Trichoptera – 9,8 тыс. экз/ м² и у личинок Сhironomidae – 15,1 тыс. экз/ м² в час (рис. 4).
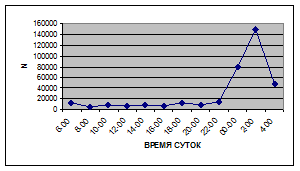
Иногда максимальная интенсивность дрифта в течение суток достигает показателей в 148,7 тыс. экз/ м² в час (18 июля 2006 г.), а в другие дни она может быть ниже в несколько раз (6 июля 2005 г. – 23,8 тыс. экз/ м² в час).
Вероятно, различия показателей интенсивности дрифта напрямую связаны с погодными и гидрологическими условиями. Так, 6 июля 2005 г. температура воздуха была +13…+150С, а 18 июля 2006 г. – +34…+360С, при этом уровень воды в начале июля 2005 г. был выше уровня воды в этот же период 2006 г.
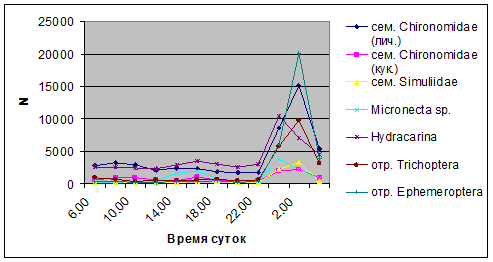
Рис. 4. Суточная динамика интенсивности массовых таксонов водной фракции дрифта (N, экз/ м² в час)
С наступлением утренних сумерек интенсивность дрифта резко уменьшается и достигает минимальных показателей – 6,5 тыс. экз/ м² в час. Возможно, к этому времени большинство организмов удовлетворили свои потребности в расселении.
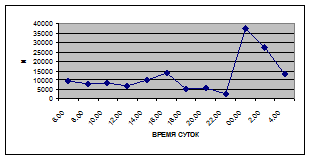
На фоне дневного минимума можно наблюдать пики второго порядка. Такие пики были отмечены 29 июня 2005 г. в шесть часов утра и в четыре часа дня, 6 июля 2005 г. – в восемь и четыре часа, 16 июля 2005 г. – в восемь и четыре часа, 18 июля 2005 г. – в четыре часа дня и 11 июля 2006 г. – в восемь и четыре часа. 19 июня, 18 и 27 июля 2006 г. пиков второго порядка вообще не наблюдалось (рис. 3).
Большую долю в максимальных пиках второго порядка составляют водные клещи Hydracarina, клопы Micronecta sp., ракообразные Corophium curvispinum Sars, 1895, личинки и куколки Сhironomidae.
Миграционная активность бентосных животных находится в противофазе с пищевой активностью хариуса – одного из наиболее многочисленных видов рыб на Сылве (Паньков, 2002). Низкие показатели дрифта утром можно объяснить наибольшей активностью хариуса в эти часы.
В целом показатели интенсивности дрифта р. Сылвы имеют довольно высокие значения и сильно отличаются от литературных данных по разным водотокам. Так для р. Лувеньги (Кольский полуостров) интенсивность дневного дрифта изменялась от 45 до 125 экз/ м² в час в конце июня – начале июля (Задорина, 1987б); для р. Лижмы (бассейн Онежского озера) интенсивность дрифта составила 554–1545 экз/ м² в час (Шустов, Широков, 1980); в р. Порье (бассейн Белого моря) интенсивность дрифта в июле составила 665–1194 экз/ м² в час (Задорина, 1987а).