Основные закономерности передачи наследственных свойств
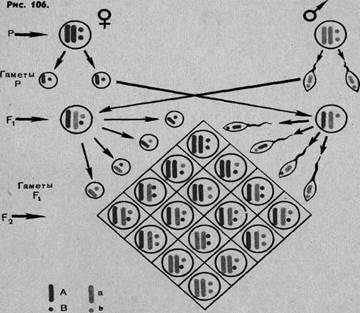
Рис. 2. Цитологические основы дигибридного скрещивания. Хромосомы, несущие доминантные гены, красные, рецессивные – синие.
Зная, что аллельные гены локализованы в гомологичных хромосомах, мы можем несколько иначе изобразить ход дигибридного скрещивания и расщепления, чем мы это делали до сих пор, представив в формулах гомологичные хромосомы в виде черточек.
Анализирующее скрещивание. Все изложенное выше о закономерностях в характере наследования признаков ясно показывает, что по фенотипу организма нельзя судить с достаточной полнотой о его наследственной структуре — его генотипе. Например, горох с желтыми гладкими семенами может быть гомозиготным (генотип ААВВ), а может быть и дигетерозиготным (АаВb) или гетерозиготным по одной аллели (ААВb и АаВВ). Определить генотип можно лишь по характеру расщепления гибридного поколения. Определение генотипа не только представляет теоретический интерес, но также имеет и большое практическое значение при селекционной работе по выведению или улучшению пород и сортов.
При этом широко используется анализирующее скрещивание, которое представляет собой скрещивание особи, генотип которой мы хотим определить, с формой чисто рецессивной по изучаемым аллелям. Такое скрещивание имеет большие преимущества, заключающиеся в том, что позволяют в первом же поколении гибридов определить гаметы анализируемой формы. Действительно, чисто рецессивная форма всегда гомозиготна. Например, зеленые морщинистые семена гороха имеют генотип ааbb и дают гаметы только одного вида — ab. Допустим, что горох, обладающий зелеными морщинистыми семенами, мы скрестили с горохом с желтыми и гладкими семенами, генотип которых нам неизвестен, и получили потомство, в котором 25% растений обладают желтыми гладкими семенами, 25% — желтыми морщинистыми, 25% — зелеными гладкими и 25% — зелеными морщинистыми. Очевидно, можно утверждать, что она образовывала 4 сорта гамет в равных количествах, т. е., другими словами, была гетерозиготной по двум аллелям.
Допустим, что в другом аналогичном скрещивании растений с теми же признаками мы не получили в потомстве никакого расщепления и все растения оказались имеющими лишь доминантные признаки по изучаемым аллелям (т. е. желтые гладкие семена). Это будет указывать на то, что взятая нами особь была доминантной гомозиготной (ААВВ). Этот случай уже был разобран подробно выше (54).
2. Явление сцепленного наследования
Независимое распределение наследственных факторов (второй закон Менделя) основано на том, что гены, относящиеся к разным аллелям, размещены в разных парах гомологичных хромосом. Естественно возникает вопрос: а как же будет происходить распределение разных (неаллельных) генов в ряде поколений, если они лежат в одной и той же паре хромосом? Уже чисто теоретически следует допустить, что такое явление должно иметь место, ибо число генов, которое удается установить путем гибридологического анализа, во много раз превосходит число хромосом. Очевидно, что к генам, находящимся в одной хромосоме, закон независимого распределения (второй закон Менделя) неприложим и закон этот должен быть ограничен лишь теми случаями, когда гены разных аллелей находятся в разных хромосомах.
Вопрос о закономерностях наследования при нахождении генов в одной хромосоме был тщательно изучен Т. Морганом и его школой. Основным объектом исследований служила небольшая плодовая мушка дрозофила. Это насекомое исключительно удобно для генетической работы. Мушка легко разводится в лабораторных условиях, плодовита, каждые 20—25 дней дает новое поколение, обладает многочисленными и разнообразными наследственными изменениями, имеет небольшое число хромосом (в диплоидном наборе — 8).
Многочисленные опыты показали, что гены, локализованные в одной хромосоме, оказываются сцепленными, т. р. наследуются преимущественно вместе, не обнаруживая независимого распределения по второму закону Менделя. Рассмотрим конкретный пример. Если скрестить дрозофилу с серым телом и нормальными крыльями с мушкой, обладающей темной окраской тела и укороченными крыльями, то в первом поколении гибридов все мушки будут серыми с нормальными крыльями. При скрещивании гибридов между собой во втором поколении не произойдет независимого распределения признаков по двум аллелям («серое тело — темное тело» и «нормальные крылья — укороченные крылья») по формуле (3:1)2.
Среди особей второго поколения гораздо чаще, чем это можно было бы ожидать при независимом распределении признаков, будут встречаться мушки с серым телом и нормальными крыльями и мушки с темным телом и недоразвитыми крыльями. Лишь у очень небольшого числа мушек произойдет перекомбинация родительских признаков иполучатся мушки с серым телом и недоразвитыми крыльями и темные с нормальными крыльями. Мы видим на этом примере, что гены, обусловливающие признаки «серое тело — нормальные крылья» и «темное тело — недоразвитые крылья», наследуются преимущественно вместе, или, иначе говоря, оказываются сцепленными между собой. Это сцепление зависит от локализации генов в одной и той же хромосоме. Поэтому при мейозе эти гены не расходятся, не отделяются друг от друга, а наследуются вместе. Явление сцепления генов, локализованных в одной хромосоме, известно под названием закона Моргана.
Почему же все-таки среди гибридов второго поколения появляется небольшое число особей с перекомбинацией родительских признаков? Почему сцепление генов не является абсолютным? Исследования показали, что эта перекомбинация генов обусловлена тем, что в процессе мейоза при конъюгации гомологичных хромосом они в известном проценте случаев обмениваются своими участками, или, иначе говоря, между ними происходит перекрест. Ясно, что при этом гены, локализованные первоначально в одной хромосоме, окажутся в разных хромосомах, между ними произойдет перекомбинация. В настоящее время явление сцепления генов исследовано на многих объектах. Среди растений особенно полно изучены в этом отношении кукуруза, томаты, посевной горох.
3. Взаимодействие генов
Взаимодействие генов. Новообразования при скрещивании.
В рассмотренных выше примерах имело место относительно независимое проявление действия генов. Доминантный ген желтой окраски семян гороха вызывает развитие этого признака как в присутствии доминантного гена гладкой формы семян, так ипри наличии аллельного ему рецессивного гена морщинистой формы семян. У морских свинок ген черной или белой окраски действует независимо от генов, определяющих характер развития шерстного покрова. На основании знакомства с этими примерами может сложиться впечатление, что генотип организма слагается из суммы отдельных, независимо действующих генов. Такое представление ложно. Хотя в некоторых случаях действие генов, принадлежащих к разным аллелям, относительно независимо, но чаще между ними осуществляются разные формы взаимодействия.
Развитие того или иного признака организма обычно находится под контролем многих генов.