Отдел диатомовые водоросли
Значение пор, по-видимому, заключается в том, что через них содержимое клетки сообщается с окружающей средой.
Во многих руководствах и исследованиях можно встретиться и с несколько иной терминологией. Различают два основных элемента структуры панциря: поры и ареолы. В однослойных — ламинарных стенках образуются только поры, которые могут быть открытыми в виде сквозного отверстия или полузамкнутыми мембранами разного типа. Ареолы образуются только в двухслойных — локу-лярных стенках и обычно затянуты снаружи или внутри тонкой перфорированной пленкой—«велумом» или «крибрумом». Помимо пор, как уже указывалось, на створках имеются различною рода выросты, характер и расположение которых служат важными таксономическими признаками. Так у ряда центрофи-циевых диатомей существуют выросты: центральные, краевые, промежуточные между ними и одиночный или добавочный вырост, расположенный или между маргинальными выростами, или же между центром и краем створки. Г. Хасле подробно изучила их строение и назначение. Центральные выросты оказались полыми трубками, иногда расширенными на вершине, основание их окружено ободком в виде муфты, прерванной в четырех местах, что позволяет видеть стенки цилиндров до самого основания. Хасле назвала эти трубки «подпертыми трубками». Их функция состоит в выделении плотной слизи, образующей студенистые тяжи, соединяющие смежные клетки в це-почковидную колонию. Краевые и промежуточные выросты также трубчатые, открытые наружу и внутрь клетки, через них выделяется слизь, покрывающая всю поверхность створки. Эти выросты свойственны только планктонным видам. Крупный одиночный вырост тоже трубчатый, иногда на вершине он расширен, а внутрь клетки открывается узкой щелью с утолщенными краями в виде двух губ. Хасле назвала эти выросты двугубыми; функция их пока не ясна. Примерами строения и назначения этих выростов могут служить выросты у представителей родов талассиозира и ске-летонема {Skeletonema).
У большинства видов талассиозиры клетки образуют рыхлые цепочки посредством слизистого тяжа, соединяющего центры смежных створок. С помощью сканирующего электронного микроскопа показано, что эта соединительная нить между соседними панцирями образована несколькими нитями, которые выдавливаются из 7—12 «подпертых трубок», расположенных в центре створки. Помимо центральных подпертых трубок, на створке имеются более мелкие подпертые трубки и один двугубый отросток. У разных видов они расположены по-разному. Отличительный признак рода скелетонема {Skeletonema) — соединительные отростки— маргинальные подпертые трубки в числе 7—9 со срав-
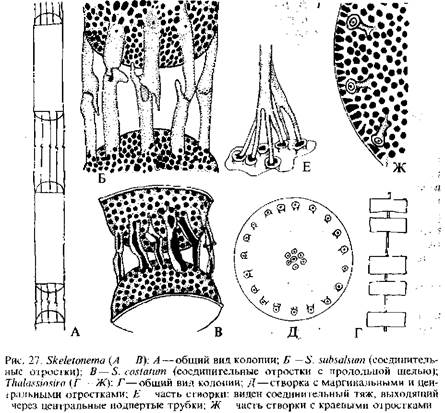
нигельно длинными наружными частями, которые смыкаются обычно по одному с такими же отростками смежных клеток, образуя рыхлые цепочки. Структура соединительных отростков у разных видов неодинакова. Так, у S. subsalsum они трубчатые, а у 5*. costatum расщеплены в виде желоба почти на протяжении всей длины.
Помимо трубковидных, створки могут нести и иные выросты, часто также служащие для объединения клеток в колонии. Например, у рода хетоцерос с цепочкообразными колониями клетки соединяются с помощью длинных негнущихся щетинок. Овальные створки биддульфии тоже снабжены выростами, которые объединяют клетки в рыхлые цепочки.
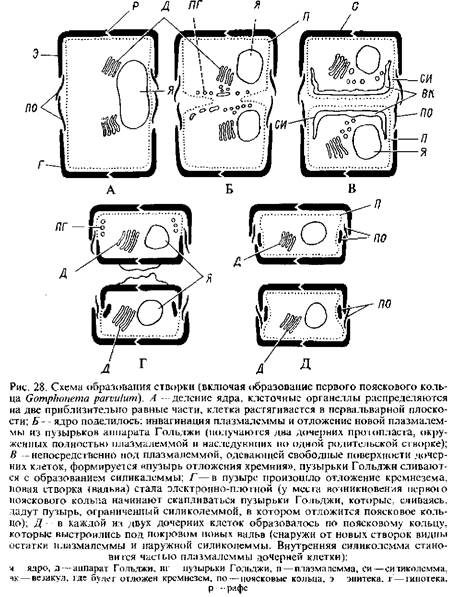
Размножение. Обычный способ размножения у диатомей — клеточное деление. В результате этого процесса оба образующихся индивида получают одну створку Т>т родительской клетки, тогда как другая возникает заново. Две створки в делящейся клетке слегка расходятся так, что остаются соединенными только края их поисковых ободков. Ядро претерпевает митоз. Расщепление протопласта начинается с периферии, идет ковнутри и всегда происходит в плоскости, параллельной створкам. На заново образовавшейся свободной поверхности обоих дочерних протопластов формируются новые створки, а затем около каждой из новых створок возникает поисковый ободок и всегда ковнутри от поисковых ободков первоначального индивида. Таким образом, поисковый ободок каждой родительской створки перекрывает поисковый ободок заново образовавшейся створки, и гипотека родительской клетки становится эпитекой одного из дочерних индивидов. Возникновение новых створок можно проследить только с помощью электронного микроскопа. Электронно-микроскопические исследования показали, что в каждой полученной в результате деления клетке, окруженной плазмалеммой, недостающаи створка формируется внутри периферического плоского, окруженного мембраной пузыря — «пузыря отложения кремнезема», который служит матрицей, определяя окончательную морфологию створки. Пузырь отложении кремнезема возникает под плазмалеммой, по-видимому, благодаря слиянию пузырьков Гольджи. Мембрана, ограничивающая этот пузырь, получила название силиколеммы; возможно, она ответственна за отложение кремнезема, т. е. за транспорт иона SiO^"2 в пузырь и его полимеризацию там до кремнезема. По достижении новой створкой зрелости и ее освобождении силиколемма, обращенная внутрь, становится плазмалеммой' и, возможно, составляет часть органического покрова, возникающего в основном за счет слияния силиколеммы и плазмалеммы, обращенных кнаружи клетки.
Ниже приводится несколько упрощенная схема процесса образования новых половинок панцири в делящейся клетке Gomphonema parvulutn. Сравнительно недавно у Fragiiaria capucina было прослежено возникновение иитеркалярных — вставочных поясков. Было показано, что они также формируются внутри пузырей отложения кремнезема, возникающих непосредственно под плазмалеммой, по-видимому, в результате слияния везикул- дериватов аппарата Гольджи. Ободки покрыты матриксом, который удерживает их вместе. Таким образом, элементы панцири являются независимыми единицами, которые и формируются в сепаратных «пузырях отложения кремнезема». Этим подтверждаются более ранние данные световой микроскопии: поясковые и вставочные ободки не образуются до тех пор, пока не будет завершено формирование створки.
В результате деления клетки одна из дочерних клеток, получившая от материнской эпитеку, точно повторяет размеры материнской клетки. Вторая дочерняя клетка, получившая от материнской гипотеку, которая стала для нее эпитекой, несколько меньших размеров: на двойную толщину пояскового ободка. Из-за окре-мнения панциря клетки могут увеличиваться только вдоль оси, соединяющей две створки. В результате размеры клеток в популяции диатомовых прогрессивно уменьшаются. Скорость этого уменьшения размеров не так уж велика, некоторое растяжение панциря у слабо окремнелых диатомей все же наблюдается и частота делений, особенно меньших из двух дочерних клеток, невелика. Специальное исследование показало, что уменьшение размеров клеток до минимальных может растягиваться на несколько лет: на два — пять у Stephanodiscus hantzschii, на четыре года у Tabellaria fenestrata. Затем следует краткий период образования растущих спор — ауксоспор, возникающих в результате полового процесса. Зигота, одетая неокремнелой растяжимой оболочкой, превращается в ауксоспору, которая растет, пока не достигнет максимальной для данного вида величины, после чего образуется панцирь и начинается ряд последовательных вегетативных делений клеток.