Отдел динофитовые водоросли
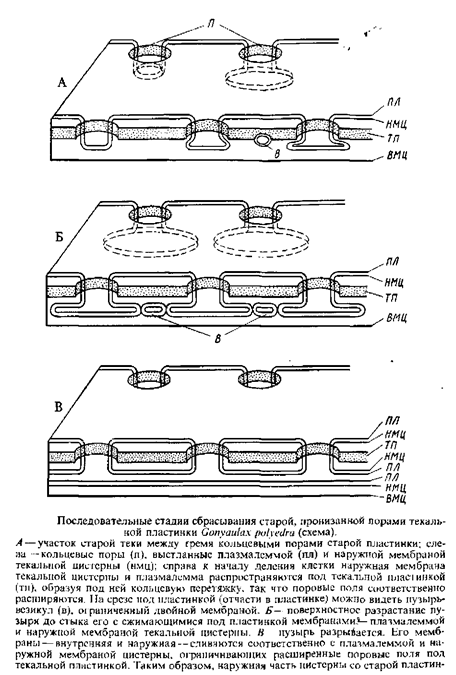

Таким образом, старые пластинки панциря сбрасываются в результате перешнуровки текальных пузырей в плоскости, параллельной поверхности. Внутренняя часть перешнурованных текальных пузырей, возможно, содержит еще старый материал для образования новых лластинок. Эти пластинки не будут отличаться от новых пластинок, возникающих в пузырях, выстилающих борозду дробления, т. е. в половинке клетки, образующей панцирь заново.
У Peridinium cinctum клетка делится внутри старого панциря, текальные пластинки которого отчасти растворяются. Дочерние клетки удерживаются некоторое время вместе внутри разрушающегося панциря, заключенные в общий фиброзный слой; после его разрыва клетки освобождаются. Еще внутри старого панциря в дочерних клепках начинается отложение текальных пластинок. Ультратонкие срезы через делящиеся клетки позволили проследить возникновение новой плазмалеммы дочерних клеток. Под внутренней мембраной текальных цистерн старого панциря появляются плоские везикулы, которые сливаются с образованием наружной и внутренней мембран. Внутренняя мембрана пузыря становится плазмалеммой дочерних клеток. Из содержимого пузыря, возможно, возникает фиброзный слой. Наружная мембрана везикула остается с внутренней мембраной текальных цистерн, старыми текальными пластинками, наружной мембраной текальных цистерн и старой плазмалеммой.
У Woloszynskia apiculata при бесполом размножении клетки теряют подвижность, округляются и превращаются в зооспорангии. При этом под текой выделяется тонкая целлюлозная оболочка собственно зооспорангия. Содержимое зооспорангия делится на две, четыре, восемь клеток, которые освобождаются при растворении целлюлозной стенки собственно зооспорангия и разрыве и сбрасывании теки исходной материнской клетки.
На протяжении двух последних десятилетий у многих дино-флагеллят рядом ученых обнаружено половое размножение, ранее известное лишь у одного из видов гленодиниума. Успех этих исследований в большой степени обусловлен развитием методов культивирования на искусственных питательных средах и накоплением данных о том, что половой процесс у многих водорослей индуцируется недостатком в среде азота. Это оказалось справедливым и для многих динофлагеллят. Так, при помещении в среду, свободную от азота, или в старых культурах, где уровень азота низок, половой процесс наблюдался у ряда видов Peridinium Woloszynskia apiculata, Glenodinium lubiensiforme, Gonyaulax moniliata, Crypthecodinium cohnii, Protogonyaulax catenella и др. Сливаются голые гаметы одинакового размера, г, е. имеет место изогамия, и зиготы у P. cinctum, P. willei, P. Umbatum, Gymnodinium pseudopalustre, Crypthecodinium cohnii возникают в пределах одного клона.
Woloszynskia apiculata изогамна, но гетероталлична, у этого вида наблюдалось образование групп. Гаметы сливаются своими боками. Напротив, у видов рода цератиум {Ceratium horridum, С. cornutum) копуляция гамет постоянно анизогамная, и сам процесс слияния иной: мужская гамета входит в женскую через отверстие поблизости от места прикрепления жгутиков.
Отличительной чертой всех до сих по"р изученных в этом отношении динофлагеллят является долго продолжающийся рост планозиготы. Так, планозиготы Gymnodinium pseudopalustre, прежде чем превратиться в покоящуюся зиготу, растут 11 дней при 15'' С или 8 дней при 21 ° С. У Woloszynskia apiculata планозиготы также сильно вырастают и остаются подвижными в течение 8—10 или 3—4 дней при температурах 15 и 21° С соответственно. Планозиготы передвигаются с помощью двух сохраняющихся задних жгутиков гамет, в некоторых случаях остаются и поперечные жгутики, по крайней мере один из них. Динофлагелляты с «двойными» задними жгутиками описывались более 100 лет тому назад и в материале из природы: по-видимому, в этих случаях речь идет о планозиготах, которые нередко таким образом возникают и в природных условиях, а не только в культурах. После длительного движения, сопровождающегося ростом, планозиготы превращаются в неподвижные толстостенные гипнозиготы.
Г. Дюр исследовал строение оболочек гипноспор в электронном микроскопе. Наружная оболочка похожа на панцирь вегетативных клеток, отличаясь от него большей толщиной пластинок и более широкими швами. Образование средней и внутренней оболочек, по-видимому, сходно с образованием теки, т. е. они состоят из пластинок, которые возникают в текальных цистернах, однако швы отсутствуют. Это заставляет предположить, что цистерны, в которых образуются пластинки мезоспория и соответственно эндоспория, сливаются с образованием непрерывных наружной и внутренней мембран: затем между ними возникают мезоспорий и эндоспорий.
Через некоторый период покоя зиготы прорастают: так, отдельные зиготы Gymnodinium pseudopalustre и Woloszynskia apiculata способны прорастать через несколько недель на свету при 15" С. Однако более обильное и синхронизированное прорастание бывает при охлаждении зигот по крайней мере в течение 4 недель при 6 С для Woloszynskia и при 3е С для Gymnodinium и последующем переносе их на свет в условия более высокой температуры. При прорастании зигот всех изученных до сих пор динофлагеллят происходит мейоз, и таким образом динофлагелляты -—гаплонты, т. е. их ядра в вегетативных клетках имеют гаплоидный набор хромосом.
* * *
Упомянутые выше роды динофлагеллят распределяются по многим семействам. Кроме них можно назвать еще род ночесветка {Noctiluca) и ряд родов, объединяемых в семейство динофизовые, заметно отличающиеся от вышеописанных динофлагеллят.
Род ночесветка — крупная, распространенная по всему свету морская бесцветная голозойная динофлагеллята. Клетка сферическая, достигающая 1 - 2 мм в диаметре, покрыта снаружи полиэдрическими, одетыми мембраной текальными везикулами, снабженными трихоцистными порами. Изнутри везикулы выстланы рядами микротрубочек, тянущихся параллельно поверхности клетки. Цитоплазма образует гонкий периферический слой, от которого отходят тонкие тяжи, пересекающие крупные вакуоли с клеточным соком, Постенная цитоплазма содержит многочисленные липидные везикулы; некоторые из них, возможно, являются «микроисточниками» биолюминесценции. На одной стороне сферической клетки имеется глотка, в которой расположен один короткий жгутик. Из этой части клетки образуется крупное поперечно исчерченное подвижное щупальце, функция которого — улавливать необходимые для питания организмы в свой слизистый эксудат и увлекать их в глотку, где они заключаются в пищеварительные вакуоли. В цитоплазме находятся ядро и крупные митохондрии, трихоцисты. По соседству с местом прикрепления щупальца расположены пучки исчерченных волокон, возможно, контролирующих его движение.
Наряду с делением — наиболее обычным способом размножения у ночесветки описаны: образование многих зооспор, похожих на гимнодиниум, и половой процесс в.виде слияния од-ножгутиковых изогамет, возникающих внутри материнской клетки в числе 1024. Есть данные, что перед образованием гамет происходит мейоз, за которым следуют многие синхронные митозы. Если эти данные подтвердятся, то ночесветка будет представлять исключение среди других динофлагеллят, у которых редукция зи-готическая.