Роль материнского генома в развитии потомка
Перенос белков через мембраны митохондрий и хлоропластов в принципе аналогичен переносу их через мембраны эндоплазматического ретикулума. Однако здесь есть несколько важных отличий. Во-первых, при транспорте в матрикс или строму белок проходит как через наружную, так и через внутреннюю мембрану органеллы, тогда как при переносе в просвет эндоплазматического ретикулума молекулы проходят только через одну мембрану. Кроме того, перенос белков в ретикулум осуществляется с помощью механизма направленного выведения (vectorial discharge)-он начинается тогда, когда белок еще не полностью сошел с рибосомы (котрансляционный импорт), а перенос в митохондрии и хлоропласты происходит уже после того, как синтез белковой молекулы будет полностью завершен (посттрансляционный импорт).
Несмотря на эти различия, и в том и в другом случае клетка синтезирует белки-предшественники, содержащие сигнальную последовательность, которая определяет, к какой мембране направится данный белок. По-видимому, во многих случаях эта последовательность отщепляется от молекулы-предшественника после завершения транспортного процесса. Однако некоторые белки сразу синтезируются в окончательном виде. Полагают, что в таких случаях сигнальная последовательность заключена в полипептидной цепи готового белка. Сигнальные последовательности еще плохо изучены, но, вероятно, должно быть несколько типов таких последовательностей, каждый из которых определяет перенос белковой молекулы в определенную область клетки. Например, в растительной клетке некоторые из белков, синтез которых начинается в цитозоле, транспортируются затем в митохондрии, другие - в хлоропласты, третьи - в пероксисомы, четвертые - в эндоплазматический ретикулум. Сложные процессы, приводящие к правильному внутриклеточному распределению белков, только сейчас становятся понятными.
Помимо нуклеиновых кислот и белков для построения новых митохондрий нужны липиды. В отличие от хлоропластов митохондрии получают бóльшую часть своих липидов извне. В животных клетках фосфолипиды, синтезированные в эндоплазматическом ретикулуме, транспортируются к наружной мембране митохондрий с помощью особых белков, а затем включаются во внутреннюю мембрану; как полагают, это происходит в месте контакта двух мембран. Основная реакция биосинтеза липидов, катализируемая самими митохондриями, - это превращение фосфатидной кислоты в фосфолипид кардиолипин, который содержится главным образом во внутренней митохондриальной мембране и составляет около 20% всех ее липидов.
Размеры и форма митохондриальных геномов
К настоящему времени прочитано более 100 разных геномов митохондрий. На-бор и количество их генов в митохондриальных ДНК, для которых полностью определена последовательность нуклеотидов, сильно различаются у разных ви-дов животных, растений, грибов и простейших. Наибольшее количество генов обнаружено в митохондриальном геноме жгутикового простейшего Rectinomo-nas americana — 97 генов, включая все кодирующие белок гены, найденные в мтДНК других организмов. У большинства высших животных геном митохон-дрий содержит 37 генов: 13 для белков дыхательной цепи, 22 для тРНК и два для рРНК (для большой субъединицы рибосом 16S рРНК и для малой 12S рРНК). У растений и простейших, в отличие от животных и большинства гри-бов, в митохондриальном геноме закодированы и некоторые белки, входящие в состав рибосом этих органелл. Ключевые ферменты матричного полинуклеоти-дного синтеза, такие как ДНК-полимераза (осуществляющая репликацию мито-хондриальной ДНК) и РНК-полимераза (транскрибирующая геном митохон-дрий), зашифрованы в ядре и синтезируются на рибосомах цитоплазмы. Этот факт указывает на относительность автономии митохондрий в сложной иерар-хии эвкариотической клетки.
Геномы митохондрий разных видов отличаются не только по набору ге-нов, порядку их расположения и экспрессии, но по размеру и форме ДНК. По-давляющее большинство описанных сегодня митохондриальных геномов пред-ставляет собой кольцевые суперспирализованные двуцепочечные молекулы ДНК. У некоторых растений наряду с кольцевыми формами имеются и линей-ные, а у некоторых простейших, например инфузорий, в митохондриях обнару-жены только линейные ДНК.
Как правило, в каждой митохондрии содержится несколько копий ее ге-нома. Так, в клетках печени человека около 2 тыс. митохондрий, и в каждой из них — по 10 одинаковых геномов. В фибробластах мыши 500 митохондрий, со-держащих по два генома, а в клетках дрожжей S.cerevisiae — до 22 митохон-дрий, имеющих по четыре генома.
Митохондриальный геном растений, как правило, состоит из нескольких молекул разного размера. Одна из них, “основная хромосома”, содержит боль-шую часть генов, а кольцевые формы меньшей длины, находящиеся в динами-ческом равновесии как между собой, так и с основной хромосомой, образуются в результате внутри- и межмолекулярной рекомбинации благодаря наличию по-вторенных последовательностей (рис.1).
Рис 1. Схема образования кольцевых молекул ДНК разного размера в митохондриях растений. Рекомбинация происходит по повторенным участкам (обозначены синим цветом).
В митохондриях большинства организмов (кроме высших животных) часть кольцевых молекул ДНК присутствует в виде олигоме-ров, которые можно разделить на три класса: линейные; кольцевые, имеющие контурную длину, кратную длине мономерных колец; цепные, катенаны, состо-ящие из топологически связанных, т.е. продетых друг в друга, мономерных ко-лец (рис.2). Так, в единственной митохондрии простейших из отряда кинето-пластид, включающего эндопаразита человека — трипаносому, содержатся ты-сячи кольцевых молекул ДНК. У Trypanosoma brucei имеются два типа моле-кул: 45 одинаковых максиколец, каждое из которых состоит из 21 тыс. пар ну-клеотидов, и 5.5 тыс. идентичных друг другу миниколец по 1000 пар нуклео-тидов. Все они, соединяясь в катенаны, образуют переплетенную сеть, которая вместе с белками формирует структуру, называемую кинетопластом.
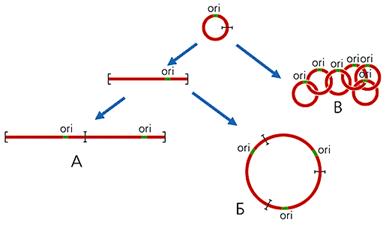 Рис 2. Схема образования линейных (А), кольцевых (Б), цепных (В) олигомеров мтДНК. ori — район начала репликации ДНК.
Рис 2. Схема образования линейных (А), кольцевых (Б), цепных (В) олигомеров мтДНК. ori — район начала репликации ДНК.
Размер генома митохондрий разных организмов колеблется от менее 6 тыс. пар нуклеотидов у малярийного плазмодия (в нем, помимо двух генов рРНК, содержится только три гена, кодирующих белки) до сотен тысяч пар ну-клеотидов у наземных растений (например, у Arabidopsis thaliana из семейства крестоцветных 366924 пар нуклеотидов). При этом 7—8-кратные различия в ра-змерах мтДНК высших растений обнаруживаются даже в пределах одного се-мейства. Длина мтДНК позвоночных животных отличается незначительно: у человека — 16569 пар нуклеотидов, у свиньи — 16350, у дельфина — 16330, у шпорцевой лягушки Xenopus laevis — 17533, у карпа — 16400. Эти геномы схо-дны также и по локализации генов, большинство которых располагаются встык; в ряде случаев они даже перекрываются, обычно на один нуклеотид, так что по-следний нуклеотид одного гена оказывается первым в следующем. В отличие от позвоночных, у растений, грибов и простейших мтДНК содержат до 80% не-кодирующих последовательностей. У разных видов порядок генов в геномах митохондрий отличается.