Свет и его роль в жизни растений и животных
5. Гамма-аминомасляная кислота
Одним из главных компонентов пула свободных аминокислот головного мозга различных животных является у-аминомасляная кислота, продукт а-декарбоксилирования глута-миновой кислоты. Цикл превращений ГАМК в мозге включает три сопряженных энзиматические реакции, получившие название ГАМК-шунта.
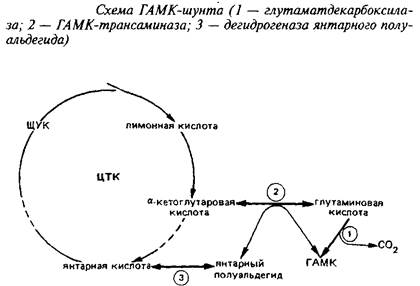
Он является ответвлением ЦТК на участке от а-кетоглутарата до сукцината. При участии фермента i лугам атдекарбокс ил азы отщепляется первый карбоксил L-глутаминовой кислоты с образованием ГАМК.
Этот энзим присутствует только в ЦНС и главным образом в сером веществе. ГДК синтезируется в нейрональной соме, а затем очень быстро транспортируется вдоль аксона. ГДК нуждается в пиридоксальфосфате в качестве кофактора, как большинство других декарбоксилаз аминокислот. Кофактор прочно связан с энзимом. Молекулярная масса энзима 85 кД, Kw для глутамата около 0,7 мМ, а Км для пиридоксальфосфата 0,05 М. ГДК специфичен для глутамата, слабо взаимодействует с аспарагиновой кислотой. Скорость ГДК-реакции - лимитирующая ступень синтеза ГАМК. Уровень ГАМК в различных областях нервной системы регулируется действием ГДК и при нормальных условиях мало зависит от действия энзимов деградации ГАМК. ГДК является маркером ГАМК-ергических синапсов.
Энзимы катаболизма ГАМК локализованы отдельно от ГДК. ГАМК-трансаминаза находится в сером веществе мозга, но встречается также и в других тканях. Она также требует пиридоксальфосфат в качестве кофактора и связана с ним прочно. ГАМК-Т обнаружена в митохондриях, в то время как ГДК и ГАМК локализованы в синаптосомах. Км ГАМК-Т для всех субстратов очень высока.
Конечный энзим шунта - дегидрогеназа янтарного полуальдегида - превращает янтарный полуальдегид в янтарную кислоту. Он распространен в ЦНС там же, где и ГАМК-Т. Это митохондриальный энзим, который специфичен для янтарного полуальдегида и НАД"", активируется сульфгидрнлъными реагентами и подавляется субстратом при концентрации последнего выше 10~М.
ГАМК является наиболее широко распространенным медиатором торможения в нервной системе. У млекопитающих она локализована в нервных окончаниях тормозных нейронов ЦНС. ГАМК тормозит биоэлектрическую активность не только головного мозга позвоночных, но и нервных цепочек и ганглиев беспозвоночных животных. Соответственно ГАМК и ферменты ее обмена также локализованы в нервных структурах беспозвоночных, совпадающих с расположением тормозных синапсов. Физиологическое действие ГАМК обусловлено взаимодействием со специальными рецепторами.
6. Компартментализация метаболизма аминокислот
Компартментализация метаболизма является ключевым фактором взаимоотношений между глутаматом, глутамином и ГАМК, Впервые это явление было открыто в лаборатории Вэлша в конце 50-х - начале 60-х годов и известно под названием эффекта Вэлша. При определенных условиях в опытах с использованием меченых предшественников специфическая радиоактивность продукта, образованного в короткий промежуток времени, превышает специфическую активность предшественника иногда в несколько раз. Эти наблюдения позволяют сделать заключение, что метаболизм имеет место в малом, высокоактивном пуле, а меченый предшественник разбавляется при выделении большим количеством немеченого предшественника из другого, малоактивного пула.
Инъекция меченого глутамата, аммония, бикарбоната, ацетата, бутирата, цитрата и других, как правило, приводила к тому, что специфическая радиоактивность глутамина была выше предшественника, изолированного вскоре после инъекции. Этот эффект не был обнаружен после инъекции меченой глюкозы, пирувата, лактата, глицерина. Данные позволили заключить, что глюкогенные субстраты метаболируют до аминокислот в компартментах, отличных от тех, в которых обмениваются кетогенные субстраты.
Эффект Вэлша - специфическое свойство нервной системы и демонстрируется в опытах как in vitro, так и in vivo. В дальнейшем кинетическими исследованиями с различными метаболическими предшественниками было показано наличие в головном мозге различных метаболических компартментов цикла трикарбоновых кислот и аминокислот, связанных с этим циклом. Некоторые исследователи ограничивают число компартментов двумя - большим и малым, другие описывают до шести метаболических компартментов. Очевидным является факт, что каждый компартмент является суммой большого числа микрокомпартментов с более или менее сходными метаболическими свойствами.
"Большой" компартмент включает в себя относительно большие пулы промежуточных соединений, которые быстро обменивается с большим пулом глутамата и малым пулом глутамина. Глюкоза используется во всех компартментах, но в большой компартмент включается до 90% гликолитического потока и большая часть общего потока через ЦТК. Глюкогенные предшественники метаболируют преимущественно в этом ком-партменте, и глюкоза может рассматриваться как предпочтительный метаболит большого компартмента. Этот же компартмент содержит основную часть общего глутамата и аспартата. Однако скорость синтеза глутамина в нем относительно низка.
Полагают, что "большой" компартмент расположен главным образом в нейронах и участвует преимущественно в энергетических процессах. Предположение о нейрональной локализации "большого" метаболического компартмента подтверждается, например, исследованиями на животных с различным ти-реоидным статусом. Так, удаление щитовидной железы при рождении животного выливается в недоразвитие нейрональных систем. Одновременно наблюдается недоразвитие "большого" метаболического компартмента. Напротив, обработка тиреоид-ными гормонами ускоряет созревание мозга и развитие метаболической компартментации.
"Малый" метаболический компартмент включает в себя ЦТК, но с малыми пулами его компонентов, которые быстро обмениваются с малым пулом глутамата, находящимся, в свою очередь, в равновесии с большим пулом глутамина. "Малый" компартмент является главным источником глутамина. Окислительная способность "малого" компартмента низка; вероятно, он не богат структурами, вовлекаемыми в синтез белка, и митохондриями. Морфологическая характеристика астроглии соответствует биохимическим свойствам "малого" метаболического компартмента. Пул глутамата, связанный с синтезом глутамина, составляющий малую долю общего пула глутамата, находится в астропи-тах. Последние составляют лишь около четверти от общего объема ткани мозга, причем концентрация глутамата в них ниже, чем в ткани ЦНС в целом. Коммуникация между "малым" и "большими" компартментами осуществляется через транспорт глутамина и ГАМК, а также путем аксонального тока белков из нейронального перикариона к нервным окончаниям.
Значение метаболических компартментов состоит в пространственном отделении биосинтетических процессов от тех метаболических путей, которые строго контролируются энергетическими нуждами. Это явление характерно именно для нервной ткани, которая отличается большой функциональной гетерогенностью составляющих ее элементов, большой долей крупных и средних клеток с разнообразными системами органелл и, наконец, большой протяженностью отростков нервных клеток, что затрудняет возможность смешивания метаболитов.