Физиология зрительного и слухового анализатора
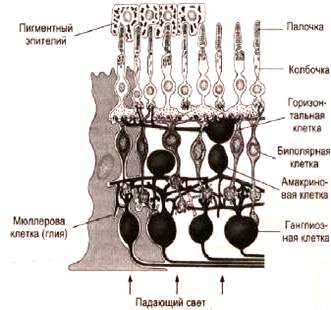
Аксоны биполярных клеток в свою очередь конвергируют на ганглиозные клетки (второй нейрон). В результате на каждую ганглиозную клетку могут конвергировать около 140 палочек и 6 колбочек, при этом чем ближе к желтому пятну, тем меньше фоторецепторов конвергирует на одну клетку. В области желтого пятна конвергенция почти не осуществляется и количество колбочек почти равно количеству биполярных и ганглиозных клеток. Именно это объясняет высокую остроту зрения в центральных отделах сетчатки.
Периферия сетчатки отличается большой чувствительностью к слабому свету. Это обусловлено, по-видимому, тем, что до 600 палочек конвергируют здесь через биполярные клетки на одну и туже ганглиозную клетку. В результате сигналы от множества палочек суммируются и вызывают более интенсивную стимуляцию этих клеток.
В ганглиозных клетках даже при полном затемнении спонтанно генерируются серии импульсов с частотой 5 в секунду. Эта импульсация обнаруживается при микроэлектродном исследовании одиночных зрительных волокон или одиночных ганглиозных клеток, а в темноте воспринимается как "собственный свет глаз".
В одних ганглиозных клетках учащение фоновых разрядов происходит на включение света (оn-ответ), в других - на выключение света (off-ответ). Реакция ганглиозной клетки может быть обусловлена и спектральным составом света.
В сетчатке кроме вертикальных существуют также латеральные связи. Латеральное взаимодействие рецепторов осуществляется горизонтальными клетками. Биполярные и ганглиозные клетки взаимодействуют между собой за счет многочисленных латеральных связей, образованных коллатералями дендритов и аксонов самих клеток, а также с помощью амакриновых клеток.
Горизонтальные клетки сетчатки обеспечивают регуляцию передачи импульсов между фоторецепторами и биполярами, регуляцию цветовосприятия и адаптации глаза к различной освещенности. В течение всего периода освещения горизонтальные клетки генерируют положительный потенциал - медленную гиперполяризацию, названную S-потенциалом (от англ. slow - медленный). По характеру восприятия световых раздражений горизонтальные клетки делят на два типа:
1) L-тип, в котором S-потенциал возникает при действии любой волны видимого света;
2) С-тип, или "цветовой", тип, в котором знак отклонения потенциала зависит от длины волны. Так, красный свет может вызвать их деполяризацию, а синий - гиперполяризацию. Полагают, что сигналы горизонтальных клеток передаются в электротонической форме.
Горизонтальные, а также амакриновые клетки называют тормозными нейронами, так как они обеспечивают латеральное торможение между биполярными или ганглиозными клетками.
Совокупность фоторецепторов, посылающих свои сигналы к одной ганглиозной клетке, образует ее рецептивное поле. Вблизи желтого пятна эти поля имеют диаметр 7 - 200 нм, а на периферии - 400 - 700 нм, т.е. в центре сетчатки рецептивные поля маленькие, а на периферии сетчатки они значительно больше по диаметру. Рецептивные поля сетчатки имеют округлую форму, построены концентрически, каждое из них имеет возбудительный центр и тормозную периферическую зону в виде кольца. Различают рецептивные поля с on-центром (возбуждаются при освещении центра) и с off-центром (возбуждаются при затемнении центра). Тормозная кайма, как предполагают в настоящее время, образуется горизонтальными клетками сетчатки по механизму латерального торможения, т.е. чем сильнее возбужден центр рецептивного поля, тем большее тормозное влияние он оказывает на периферию. Благодаря таким типам рецептивных полей (РП) ганглиозных клеток (с on - и off-центрами) происходит обнаружение светлых и темных объектов в поле зрения уже на уровне сетчатки.
При наличии у животных цветового зрения выделяют цветооппонентную организацию РП ганглиозных клеток сетчатки. Эта организация состоит в том, что определенная ганглиозная клетка получает возбуждающие и тормозные сигналы от колбочек, имеющих разную спектральную чувствительность. Например, если "красные" колбочки оказывают возбуждающее действие на данную ганглиозную клетку, то "синие" колбочки ее затормаживают. Обнаружены разные комбинации возбуждающих и тормозящих входов от разных классов колбочек. Значительная часть цветооппонентных ганглиозных клеток связаны со всеми тремя типами колбочек. Благодаря такой организации РП отдельные ганглиозные клетки становятся избирательными к освещению определенного спектрального состава. Так, если возбуждение возникает от "красных" колбочек, то возбуждение сине - и зеленочувствительных колбочек вызовет торможение этих клеток, а если ганглиозная клетка возбуждается от синечувствительных колбочек, то она тормозится от зелено - и красночувствительных и т.д.
Центр и периферия рецептивного поля имеют максимальную чувствительность в противоположных концах спектра. Так, если центр рецептивного поля отвечает изменением активности на включение красного света, то периферия аналогичной реакцией отвечает на включение синего. Ряд ганглиозных клеток сетчатки имеет так называемую дирекционную чувствительность. Она проявляется в том, что при движении стимула в одном направлении (оптимальном) ганглиозная клетка активируется, при другом направлении движения - реакция отсутствует. Предполагают, что избирательность реакций этих клеток на движение в разных направлениях создается горизонтальными клетками, имеющими вытянутые отростки (теледендриты), с помощью которых направленно тормозятся ганглиозные клетки. Вследствие конвергенции и латеральных взаимодействий рецептивные поля соседних ганглиозных клеток перекрываются. Это обусловливает возможность суммации эффектов световых воздействий и возникновение взаимных тормозных отношений в сетчатке.
Электрические явления в сетчатке. В сетчатке глаза, где локализуется рецепторный отдел зрительного анализатора и начинается проводниковый отдел, в ответ на действие света происходят сложные электрохимические процессы, которые можно зарегистрировать в виде суммарного ответа - электроретинограммы (ЭРГ).

ЭРГ отражает такие свойства светового раздражителя, как цвет, интенсивность и длительность его действия. ЭРГ может быть зарегистрирована от целого глаза или непосредственно от сетчатки. Для ее получения один электрод помещают на поверхность роговой оболочки, а другой прикладывают к коже лица вблизи глаза или на мочку уха.
На ЭРГ, зарегистрированной при освещении глаза, различают несколько характерных волн. Первая негативная волна, а представляет собой небольшое по амплитуде электрическое колебание, отражающее возбуждение фоторецепторов и горизонтальных клеток. Она быстро переходит в крутонарастающую позитивную волну b, которая возникает в результате возбуждения биполярных и амакриновых клеток. После волны b наблюдается медленная электроположительная волна с - результат возбуждения клеток пигментного эпителия. С моментом прекращения светового раздражения связывают появление электроположительной волны d. Показатели ЭРГ широко используются в клинике глазных болезней для диагностики и контроля лечения различных заболеваний глаза, связанных с поражением сетчатки.