Физиология мышц
Строение мышцы
Скелетная мышца состоит из пучков вытянутых в длину клеток - мышечных волокон, обладающих тремя свойствами: возбудимостью, проводимостью и сократимостью. Отличительной чертой мышечных клеток от клеток, не обладающих свойством сократимости, является наличие саркоплазматического ретикулума. Он представляет собой замкнутую систему внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу. В мембране саркоплазматического ретикулума находятся две транспортные системы, обеспечивающие освобождение от ретикулума ионов кальция при возбуждении и их возврат из миоплазмы обратно в ретикулум при расслаблении мышцы. В механизме освобождения ионов кальция из ретикулума при возбуждении мышечной клетки важную роль играет система поперечных трубочек (Т-система), представляющих собой выпячивания поверхностной мембраны мышечного волокна.
Мышечные волокна имеют диаметр от 10 до 100 мкм и длину от 5 до 400 мм (в зависимости от длины мышцы). В каждом мышечном волокне содержится до 1000 и более сократительных элементов миофибрилл, толщиной 1-3 мкм. Каждая миофибрилла состоит из множества параллельно лежащих толстых и тонких нитей - миофиламентов. Толстые нити состоят из молекул белка миозина, а тонкие - из белка актина.
Расположение миозиновых и тонких актиновых белковых нитей строга упорядочено (рис.4.1). Пучок лежащих в середине саркомера нитей миозина выглядит в световом микроскопе как темная полоска. Благодаря свойству двойного лучепреломления в поляризованном свете (то есть анизотропии) она называется А-диском. По обе стороны от А-диска находятся участки, которые содержат только тонкие нити актина и поэтому выглядят светлыми. Эти изотропные J-диски тянутся до Z-пластин. Благодаря такому периодическому чередованию светлых и темных полос миофибриллы скелетной мышцы выглядят исчерченными (поперечно - полосатыми). Если мышца расслаблена, то в средней части А-диска различается менее плотная Н-зона, состоящая только из толстых миофиламентов. Н-зона не просматривается во время сокращения мышцы. По середине J-диска проходит темная полоска - это Z линия. Участок миофибриллы между двумя Z линиями называется саркомером.
Схема саркомера мышечного волокна и взаимного расположения толстых миозиновых и тонких актиновых миофиламентов.
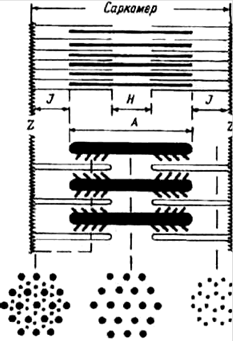
Z - линии, разделяющие два соседних саркомера; J - изотропный диск; А - анизотропный диск; Н - участок с уменьшенной анизотропностью
Механизмы сокращения мышечного волокна. В покоящихся мышечных волокнах при отсутствии импульсации мотонейрона поперечные миозиновые мостики не прикреплены к актиновым миофиламентам.
При сокращении мышцы длина А-дисков не меняется, J - диски укорачиваются, а Н-зона А-дисков может исчезать (рис.4.3). Эти данные явились основой для создания теории, объясняющей сокращение мышцы механизмом скольжения (теорией скольжения) тонких актиновых миофиламентов вдоль толстых миозиновых. В результате этого миозиновые миофиламенты втягиваются между окружающими их актиновыми. Это приводит к укорочению каждого саркомера, а значит, и всего мышечного волокна.
Молекулярный механизм сокращения мышечного волокна состоит в том, что возникающий в области концевой пластинки потенциал действия распространяется по системе поперечных трубочек вглубь волокна, вызывает деполяризацию мембран цистерн саркоплазматического ретикулума и освобождение из них ионов кальция. Свободные ионы кальция в межфибриллярном пространстве запускают процесс сокращения. Совокупность процессов, обуславливающих распространение потенциала действия вглубь мышечного волокна, выход ионов кальция их саркоплазматического ретикулума, взаимодействие сократительных белков и укорочение мышечного волокна называют "электромеханическим сопряжением". Временная последовательность между возникновением потенциала действия мышечного волокна, поступлением ионов кальция к миофибриллам и развитием сокращения волокна показана на рисунке.
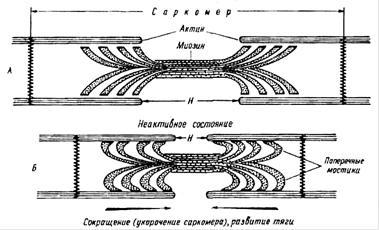
Схема сокращения мышцы
А. Поперечные мостики между актином и миозином разомкнуты. Мышца находится в расслабленном состоянии. Б. Замыкание поперечных мостиков между актином и миозином. Совершение головками мостиков гребковых движений по направлению к центру саркомера. Скольжение актиновых нитей вдоль миозиновых, укорочение саркомера, развитие тяги.
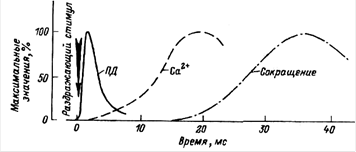
Схема временной последовательности развития возбуждения и сокращения мышцы
При возбуждении волокна Са+2 выходит из цистерн саркоплазматического ретикулума и, следовательно, концентрация его вблизи миофибрилл возрастает. Под влиянием активирующих ионов Са молекула тропонина изменяет свою форму таким образом, что выталкивает тропомиозин в желобок между двумя нитями актина, освобождая тем самым участки для прикрепления миозиновых поперечных мостиков к актину. В результате поперечные мостики прикрепляются к актиновым нитям. Поскольку головки миозина совершают "гребковые" движения в сторону центра саркомера происходит "втягивание" актиновых миофиламентов в промежутки между толстыми миозиновыми нитями и укорочение мышцы.
Источником энергии для сокращения мышечных волокон служит АТФ.
При однократном движении поперечных мостиков вдоль актиновых нитей (гребковых движениях) саркомер укорачивается примерно на 1% его длины. Следовательно, для полного изотонического сокращения мышцы необходимо совершить около 50 таких гребковых движений. Только ритмическое прикрепление и отсоединение головок миозина может втянуть нити актина вдоль миозиновых и совершить требуемое укорочение целой мышцы. Напряжение, развиваемое мышечным волокном, зависит от числа одновременно замкнутых поперечных мостиков. Скорость развития напряжения или укорочения волокна определяется частотой замыкания поперечных мостиков, образуемых в единицу времени, то есть скоростью их прикрепления к актиновым миофиламентам. С увеличением скорости укорочения мышцы число одновременно прикрепленных поперечных мостиков в каждый момент времени уменьшается. Этим и можно объяснить уменьшение силы сокращения мышцы с увеличением скорости ее укорочения.
Поскольку возврат ионов кальция в цистерны саркоплазматического ретикулума идет против диффузионного градиента, то этот процесс требует затрат энергии. Ее источником служит АТФ. Одна молекула АТФ затрачивается на возврат 2-х ионов кальция из межфибриллярного пространства в цистерны. Таким образом, кальций в мышечных волокнах играет роль внутриклеточного посредника, связывающего процессы возбуждения и сокращения.
Регуляция силы сокращения мышц. Для регуляции величины напряжения мышцы центральная нервная система использует три механизма.
1. Регуляция числа активных ДЕ. Чем больше число ДЕ мышцы включается в работу, тем большее напряжение она развивает. При необходимости развития небольших усилий и соответственно малой импульсации со стороны центральных нервных структур, регулирующих произвольные движения, в работу включаются, прежде всего, медленные ДЕ, мотонейроны, которые имеют наименьший порог возбуждения. По мере усиления центральной импульсации к работе подключаются быстрые, устойчивые к утомлению ДЕ, мотонейроны которых имеют более высокий порог возбуждения. И наконец, при необходимости увеличения силы сокращения более 20-25% от максимальной произвольной силы (МП С), активируются быстрые, легко утомляемые мышечные волокна, иннервируемые крупными мотонейронами с самым высоким порогом возбуждения.