Функционирование нервных связей в зрительной системе
Уменьшение активности в зрительных путях является основным фактором, который стремится нарушить нормальное функционирование корковых нейронов. Однако клетки коры активируются не в ответ на диффузное освещение, также вызывающее активность, а на определенные контуры и формы. Для поддержания функционирования необходимы дополнительные факторы, гораздо более «тонкие» по своей природе. В частности, должно происходить взаимодействие между сигналами обоих глаз.
Бинокулярная депривация и роль конкуренции
Первым доводом против того, что уменьшение зрительной активности может привести к изменениям в работе нейронов, являются данные эксперимента, когда у детеныша обезьяны, новорожденного или рожденного путем кесарева сечения, закрывали сразу оба глаза. Исходя из предыдущего обсуждения, можно сделать предположение, что клетки в коре не будут управляться ни одним глазом. Однако, к удивлению, после бинокулярной депривации на протяжении 17 дней и дольше большинство корковых клеток выглядело полностью нормальными. Колонки ориентации имели строение, близкое к таковому у контрольной группы (рис. 1). Основным нарушением было то, что значительная часть клеток не получала бинокулярного управления (рис. 1). Кроме того, некоторые спонтанно активные клетки вообще не получали никакого управления, другим же для активности не требовался определенным образом ориентированный стимул. Тем не менее зоны коры, контролируемые обоими глазами, оставались равными в размерах, имея паттерн, аналогичный тому, что наблюдается у нормальных взрослых обезьян: в слое 4 клетки управлялись только одним глазом, четко определялись колонки при использовании авторадиографии или окраски на цитохромоксидазу. Бинокулярная депривация у котят приводила к сходным эффектам, за исключением того, что количество клеток, получающих бинокулярное управление, оставалось большим. При этом также не происходило уменьшение ветвистости аксонов ЛКТ, направляющихся в слой 4 коры. В то же самое время клетки всех слоев в пределах ЛКТ подвергались атрофии (уменьшению в размерах примерно на 40 %).
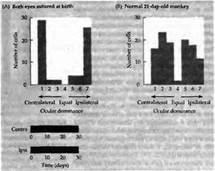
Рис. 1 Колонки зрительного доминирования после закрытия обоих глаз с рождения. (А) Обезьяна, на которой проводились исследования, была рождена посредством кесарева сечения, после чего веки обоих глаз были тотчас зашиты. Результаты были получены в возрасте 30 дней. Каждый из двух депривированных глаз все еще мог управлять клетками в зрительной коре. Рецептивные поля выглядели нормально, за исключением того, что некоторые клетки получали управление одновременно отобоих глаз. Время закрытия век показано внизу гистограммы. (В) Гистограмма глазного доминирования у нормальной обезьяны в возрасте 21 дня.
Выводом из этих экспериментов является то, что некоторые, но не все негативные эффекты, вызванные закрытием одного глаза, уменьшались или предотвращались при закрытии обоих глаз. Таким образом, импульсы от обоих глаз являются как бы конкурирующими за представление в корковых клетках, и эта конкуренция нарушается при закрытии одного глаза из двух.
Эффекты страбизма (косоглазия)
Аномалии, описанные ранее, были вызваны зашиванием век или использованием прозрачных диффузоров, приводящих к потере предметного зрения. Исходя из данных о том, что у детей, страдающих косоглазием или страбизмом (cross-eyed, strabismus, squint), а также имеющих аномальное расположение глаз (wall-eyed), также развивается слепота, Хьюбель и Визель вызывали искусственное косоглазие у котят и новорожденных обезьян, при помощи подрезания глазных мышц. В результате оптическая ось глаза отклонялась от нормальной. При этом освещение и паттерны стимуляции каждого глаза оставались без изменений.
Сначала результаты подобных опытов разочаровывали: после нескольких месяцев зрение в обоих глазах оперированного животного оставалось нормальным, и Хьюбель и Визелъ уже были готовы прекратить эту напряженную серию экспериментов (личное сообщение). Тем не менее они смогли получить следующие данные о работе корковых клеток. Отдельные корковые клетки имели нормальные рецептивные поля и живо отвечали на определенным образом ориентированные стимулы. Однако почти каждая клетка отвечала только на стимуляцию одного глаза, некоторые управлялись правым, некоторые — левым, но практически ни одна — обоими глазами. Клетки, как обычно, были сгруппированы в колонки по отношению к глазному предпочтению и к ориентации оси зрительного поля. Как и ожидалось, возникала атрофия в области ЛКТ, хотя колончатая архитектура слоя 4 оставалась без изменений. Почти полное отсутствие бинокулярного представления в коре показано на гистограмме, полученной от обезьяны с искусственно вызванным косоглазием (рис. 2). Критический период для смещенного (из-за косоглазия) восприятия, в течение которого возникают данные изменения, близок к критическому периоду монокулярной депривации.
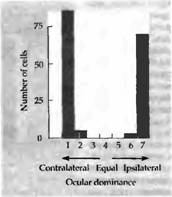
Рис. 2. Влияние косоглазия на формирование зрительного доминирования у обезьян. Глаза этой 3-летней обезьяны направлены в разные стороны, так как ей в возрасте 3 недель была проведена операция по подрезанию глазных мышц. Клетки коры получают управление либо от одного глаза, либо от другого, но не от обоих. Колонки глазного доминирования в слое 4 выглядят нормально для животного такого возраста.
Эти эксперименты представляют собой пример, когда все обычные параметры освещения в норме: количество попадающего в глаз света и паттерн стимуляции. Единственным отличием от нормы является то, что свет не падает на корреспондирующие регионы двух сетчаток. Фактором, который, скорее всего, приводит к потере бинокулярной конвергенции проекций отростков ЛКТ в кору, является недостаток соответствия в импульсах от обоих глаз. То есть гомологичные рецептивные поля обоих глаз должны быть скоординированы, чтобы возбуждение происходило одновременно. Последующие эксперименты говорят в пользу такой идеи.
В течение первых трех месяцев жизни глаза котенка были закрыты непрозрачной пластиковой пластиной, которая перемещалась с одного глаза на другой каждый день таким образом, чтобы два глаза получили одинаковый зрительный опыт, но в разное время. Результат был аналогичен данным экспериментов с искусственным косоглазием: клетки в основном управлялись либо правым, либо левым глазом, но не обоими. Поддержание нормальной бинокулярности зрения, таким образом, зависит не только от количества импульсов, но также и от надлежащего пространственного и временного соответствия активности в различных идущих к коре волокнах.
Изменения в ориентационном предпочтении
Логичным продолжением является вопрос, изменится ли ориентационное предпочтение кортикальных нейронов у животных, находящихся в условиях, когда они видят образ только одной ориентации. Экспериментальный подход, включающий в себя также конкуренцию между глазами и депривацию, был использован Карлсоном, Хьюбелем и Визелем. У новорожденной обезьяны были зашиты веки одного глаза. Животное содержалось в темноте, за исключением того времени, когда его голова помещалась в специальный держатель. В этом случае его голова фиксировалась строго вертикально и животное могло видеть вертикальные полоски оставшимся недепривированным глазом. Так как обезьяне всегда давали апельсиновый сок, когда она правильно устанавливала свою голову в держатель, она научилась быстро производить этот маневр. Таким образом, во время критического периода один глаз не получал никакой зрительной информации, а другой