Что и как закодировано в мРНК
Перенос генов из одних организмов в другие и их последующая успешная экспрессия доказали, что генетический код почти всегда универсален. Другими словами, все живые организмы от бактерий до млекопитающих пользуются в основном одним и тем же кодовым словарем. (Правда, некоторые нарушения универсальности генетического кода обнаружены в митохондриях.)
Все триплеты в транслируемой области мРНК, которые кодируют определенную аминокислоту, узнаются в рибосомах специфическими молекулами тРНК с присоединенными к ним аминокислотами. Точность узнавания весьма велика за счет существования специальной системы коррекции ошибок. Терминирующие кодоны узнаются специальными белками - факторами терминации. Надежность узнавания этих триплетов не столь высока, и иногда терминирующий триплет может прочитываться как кодон аминокислоты. Для повышения надежности процесса терминации терминирующие кодоны обычно дублируются. Первым при этом, как правило, выступает кодон UAA (основной терминирующий триплет), а вслед за ним на очень близком расстоянии в той же рамке считывания следует один из запасных терминирующих триплетов - UAG или UGA .
После расшифровки генетического кода в 1966 году вопрос о том, что и как закодировано в мРНК, казалось, был полностью исчерпан. С течением времени, однако, стало очевидно, что это далеко не так.
ПЕРЕКОДИРУЮЩИЕ СИГНАЛЫ В мРНК ИЛИ ВТОРОЙ ГЕНЕТИЧЕСКИЙ КОД
Инициирующий кодон узнается только в определенном контексте. Если мы зададим вопрос, можно ли, имея перед собой последовательность нуклеотидов какой-либо мРНК, таблицу генетического кода и зная, что трансляция мРНК идет в направлении от 5'- к 3'-концу, а белковая цепочка растет от N-конца к C-концу, написать последовательность аминокислот белка, закодированного в этой мРНК, то будем вынуждены ответить на поставленный вопрос отрицательно. Исходя из перечисленных сведений нельзя определить, с какого места мРНК мы должны начать переводить ( транслировать ) последовательность нуклеотидов в последовательность аминокислот.
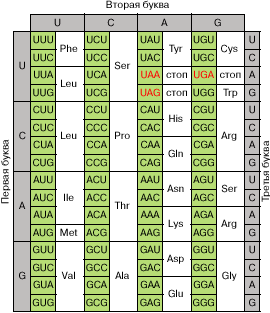
Рис. 2. Кодовый словарь. Красным цветом отмечены терминирующие кодоны
Уже очень давно стало ясно, что начало трансляции мРНК ( инициация ) не совпадает с началом самой мРНК. Свидетельством этому служат полицистронные мРНК бактерий, в которых инициация белкового синтеза происходит на каждом цистроне, а также присутствие в мРНК про- и эукариот 5'-концевых нетранслируемых последовательностей. Вместе с тем установлено, что биосинтез белка как у про-, так и у эукариот всегда начинается с одной и той же аминокислоты - метионина. Можно было бы предположить, что трансляция информации, закодированной в мРНК, начинается с первого от 5'-конца метионинового кодона, которым является триплет AUG . Для многих моноцистронных эукариотических мРНК это действительно так, хотя бывают и исключения. Однако это совсем не так для полицистронных мРНК бактерий, где инициация часто происходит на триплетах AUG, отстоящих очень далеко от начала мРНК. Этим триплетам может предшествовать большое количество других AUG, на которых инициации не происходит. Более того, оказалось, что первый метионин в белке в некоторых случаях включается не на метиониновом кодоне AUG, а на кодоне GUG , который соответствует в таблице генетического кода аминокислоте валину.
Иногда инициация с метионина может происходить и на других кодонах: AUA и AUU (кодонах изолейцина), UUG и, возможно, CUG (кодонах лейцина). Стало очевидным, что для узнавания кодона в качестве инициирующего важен не только сам и, может быть, не столько сам кодон, но какой-то контекст, делающий его инициирующим. Оказалось, что у прокариот для эффективной инициации кодон должен находиться на вершине шпилечной структуры, образуемой смежными комплементарными участками мРНК, а предшествовать ему примерно за 3-10 нуклеотидов должна полипуриновая последовательность (последовательность Шайна-Дальгарно, SD) (рис. 3, а). Эта последовательность комплементарна рибосомной РНК и, по-видимому, способствует связыванию рибосом в районе инициирующего кодона. Вторичная структура мРНК в районе инициирующего кодона и как следствие - степень доступности инициирующего кодона для инициации может изменяться в процессе трансляции соседних цистронов или при взаимодействии мРНК со специальными регуляторными белками (см. ниже).
У эукариот инициация происходит, как уже говорилось, чаще всего с первого AUG, однако только в том случае, если этот AUG находится в оптимальном контексте: за два нуклеотида до него обязательно должен находиться пурин (A или G), а непосредственно за ним должен следовать G. На эффективность инициации у эукариот определенное влияние могут оказывать также нуклеотиды и в других положениях вблизи инициирующего кодона. Самым оптимальным для узнавания инициирующего кодона у млекопитающих считается следующее его окружение: GCCGCCA / GCCAUGGA / CU (инициирующий триплет подчеркнут, а обязательные для инициации нуклеотиды выделены жирным шрифтом).
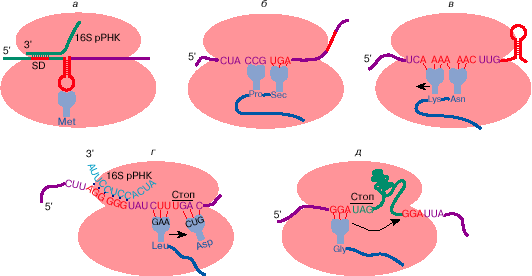
Рис. 3. Некоторые примеры отступления от общих правил трансляции генетической информации: узнавание инициирующего кодона на мРНК бактерий (а); прочитывание терминирующего кодона UGA как кодона аминокислоты селеноцистеина (б ); сдвиг рамки считывания на -1 при трансляции ретровирусной РНК (в); сдвиг рамки считывания на +1 при трансляции мРНК бактериального фактора терминации трансляции RF-2 (г); прыжок рибосомы на 50 нуклеотидов при трансляции мРНК гена 60 бактериофага Т4 (д). Рекодирующие сигналы на мРНК обозначены красным цветом. Дальнейшие пояснения см. в тексте
Если первый AUG в эукариотической мРНК находится не в оптимальном контексте, он пропускается и инициация начинается со следующего AUG. Для такой инициации очень важно также наличие кэп-структуры на 5'-конце мРНК и, как ни странно, поли(А) последовательности на противоположном конце молекулы. Кэп-структура и поли(А) последовательность узнаются специфическими белками, которые также необходимы для инициации. При таком способе инициации трансляции у эукариот последовательность мРНК как бы просматривается (сканируется) с начала мРНК (от ее кэп-структуры) для поиска кодона AUG в оптимальном контексте. Такая инициация получила название "кэп-зависимая инициация по сканирующему механизму". Следует, однако, заметить, что на некоторых мРНК эукариот инициация происходит не путем сканирования мРНК с 5'-конца, а за счет непосредственного узнавания определенного внутреннего AUG. Для такого узнавания требуется весьма протяженная предшествующая последовательность мРНК. Эта последовательность узнается особыми клеточными белками, которые способствуют инициации трансляции по механизму "внутренней инициации". По такому механизму инициируется трансляция на многих вирусных РНК, а также на некоторых клеточных мРНК, кодирующих очень важные регуляторные белки, например факторы роста фибробластов. Содержание этих белков обычно очень мало, а увеличение их количества в клетке может сопровождаться трансформацией клеток в раковые. Некоторые вирусы, генетическая информация которых считывается по механизму внутренней инициации трансляции, способны выключить инициацию трансляции клеточных мРНК по сканирующему механизму и, таким образом, переключать белоксинтезирующий аппарат клетки на синтез собственных белков.