Что такое ге. Генетическая точка зрения
Хотя в опытах Менделя исследованные признаки проявляли (к счастью) полное доминирование, так бывает не всегда. Доминирование аллелей может быть неполным (частичным) или совсем отсутствовать (что иногда называют ко доминантностью). В последнем случае гетерозиготы отличаются от гомозигот промежуточными свойствами. Например, скрещивание растений львиного зева с красными и белыми цветами дает розовые гибриды. Хотя в этом случае нет доминирования, наблюдается все та же закономерность - гибриды первого поколения (F1) одинаковы. В дальнейших скрещиваниях гибридов образуются те же численные соотношения, как и при полном доминировании, хотя вместо л чух фенотипов можно различить три.
Независимость различных генов
Во втором законе Менделя утверждается независимость комбинации различных генов. При скрещивании растения, доминантного по двум различным признакам, с растением, у которого эти признаки рецессивны, F1 по-прежнему состоит из одинаковых растений доминантного типа. Но при последующем скрещивании гибридов F1 появляются два класса растений. Один класс состоит из растений исходных родительских типов. Второй класс включает растения новых фенотипов, с доминантными признаками одного родителя и рецессивными признаками другого. Эти фенотипы называют рекомбинантными.
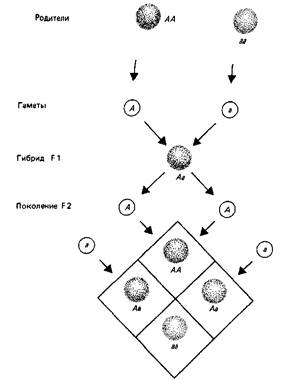
Рис.2. Аллели расщепляются независимо.
Два родителя гомозиготны: А А имеет две копии доминантного аллеля; аа имеет две копии рецессивного аллеля. Каждый образует только один тип гамет, так что F1 (первое гибридное поколение) состоит из одинаковых гибридных особей Аа. Поскольку А доминирует над а, фенотип потомков Аа такой же, как у родителя АА (показано цветом). Фенотип рецессивной гомозиготы аа показан отсутствием цвета. Каждый гибрид F1 образует гаметы А и а в одинаковом количестве. При скрещивании объединение этих гамет приведет к образованию F2 (второе гибридное поколение), которое будет состоять из \АА: 2Аа: \аа. Так как АА и Аа имеют одинаковый фенотип, получается классическое соотношение доминантного и рецессивного фенотипов-3:
1. В тех случаях, когда фенотип гетерозиготы занимает промежуточное положение между родительскими АА и аа, F1 будет другим, а в F2 отношение фенотипов будет: 1 доминантный: 2 промежуточных:! рецессивный.
На рис.3 показано, что соотношение потомков различного типа можно объяснить, предположив, что в процессе образования гамет происходит совершенно случайное объединение одного из аллелей первого признака с одним из аллелей второго. Четыре возможных типа гамет образуются в равных соотношениях; при формировании зиготы следующего поколения они объединяются случайно.
Типичное соотношение фенотипов скрывает за собой большее разнообразие генотипов, что может быть установлено путем возвратного (анализирующего) скрещивания с рецессивным родителем. По существу, возвратное скрещивание позволяет прямо установить генотип исследуемого организма (рис.4).
Закон независимого распределения генов, как следует из сказанного выше, говорит, что поведение любой пары (или большего числа) генов можно полностью предсказать с помощью правил комбинаторики. Распределение между потомками (наследование) одного гена не влияет на распределение другого. Эта концепция подразумевает, что распределение генов происходит со статистической вероятностью, а не предопределено точно. Чем больше проведено скрещиваний, тем точнее будет совпадать с предсказанным соотношение типов в потомстве.
Роль хромосом в наследственности
Открытие Менделя не сразу оценили по достоинству, поскольку не было никаких представлений о физической основе постулируемых факторов. Хромосомная теория наследственности, предложенная одновременно Саттоном и Бовери (Sutton, Boveri), разрешила долгие споры о возможной роли хромосом. Уже существовали некоторые, хотя довольно туманные, предположения о том, что хромосомы как-то участвуют в механизмах наследования. В 1903 г. наконец стало ясно, что по своим свойствам они точно совпадают с дискретными единицами наследственности, описанными Менделем.
Клеточная теория, утвердившаяся в середине девятнадцатого века, предполагала, что все организмы состоят из клеток и что эти клетки могут возникать только из предсуществующих клеток. Ранние цитологические исследования показали, что "типичная" клетка состоит из плотного ядра, отделенного мембраной, от менее плотной окружающей цитоплазмы. Внутри ядра с помощью определенных красителей можно было различить зернистые участки - хроматин. Вскоре после работы Менделя было установлено, что хроматин представлен определенным числом нитевидных образований, или хромосом.
В большинстве клеток хромосомы видны только в процессе клеточного деления. Два типа деления, присущие организмам, размножающимся половым путем, объясняют и постоянство генетического материала, и процесс наследования в том виде, как это было предсказано законами Менделя.
Клеточный цикл у растущего организма состоит из двух этапов. Более длинный период-интерфаза, когда клетка синтетически активна и занята воспроизведением своих компонентов. Затем следует короткий период-митоз, интерлюдия, во время которой фактически завершается процесс разделения на две дочерние клетки. Клетки, возникающие в результате ряда митотических делений и образующие целый организм, называют соматическими клетками.
Каждая дочерняя клетка, начинающая свою жизнь после митоза, содержит по две копии каждой хромосомы. Их называют гомологичными. Общее число хромосом в клетке, известное как диплоидный набор, обозначают 2п. Типичная соматическая клетка существует в диплоидном состоянии (кроме того периода, когда она готовится к делению или уже делится).
В интерфазе растущая клетка удваивает свой хромосомный материал. Однако это становится очевидным только в последующем митозе. В митозе каждая хромосома разделяется вдоль по длине, образуя две копии - сестринские хроматиды. В этот момент клетка содержит 4и хромосом, организованных в 2и пар сестринских хроматид. Иными словами, в клетке имеется по две (гомологичные) копии каждой пары сестринских хроматид. На рис.5 показана последовательность процессов, обеспечивающих митотическое деление. Суть заключается в том, что сестринские хроматиды растаскиваются к противоположным полюсам клетки, так что каждая дочерняя клетка получает по одной копии каждой сестринской хроматиды. Теперь это самостоятельные хромосомы.4и хромосомы, существовавшие в начале деления, разделились на два набора по 2и хромосом. Этот процесс повторяется в следующем клеточном цикле. Таким образом, митотическое деление гарантирует постоянство набора хромосом в соматических клетках.
Один родитель гомозиготен по двум доминантным генам - по гену А, определяющему цвет, и гену В, определяющему форму (круглая форма). Другой родитель гомозиготен по рецессивным аллелям а и Ъ (отсутствие цвета, морщинистая форма). Потомство F1 одинаково-с доминантным фенотипом.
Родители F1 образуют гаметы, в которых происходят независимое расщепление аллелей и независимая комбинация генов, так что образуется равное количество гамет каждого из четырех возможных типов. Они соединяются случайно и образуют 9 генотипических классов, которые из-за отношений доминантности проявляются в виде четырех фенотипических классов: 9 окрашенных гладких: 3 окрашенных морщинистых: 3 бесцветных гладких: 1 бесцветный морщинистый. Заметим, что каждый реципрокный генотип представлен одинаковым числом особей, например два родительского типа (по одному ААВВ и ааЪЬ) и два рекомбинантных (по одному каждого рекомбинантного класса ААЪЬ и ааВВ). Соотношение 3: 1 сохраняется для каждого независимо расщепляющегося признака.