Изучение вопросов биотехнологии в курсе химии средней школыРефераты >> Педагогика >> Изучение вопросов биотехнологии в курсе химии средней школы
2) Культуру протопластов на начальной стадии её роста заражают агробактериями, которые используют в качестве векторов.
3) Микроинъекции ДНК. Аналогичен методу микроинъекций животных клеток. Этот метод можно рассматривать как наиболее универсальный. Эффективность трансформации растительных клеток – 10–20% независимо от типа вектора. Трансформация не видоспецифична, возможен перенос генов в любое растение.
4) Электропорация. Метод основан на повышении проницаемости биомембран за счет действия импульсов высокого напряжения. В результате молекулы ДНК проникают в клетки через поры в клеточной мембране.
5) Упаковка в липосомы. Это один из методов, позволяющих защитить экзогенный генетический материал от разрушения нуклеазами растительной клетки. Липосомы – сферические тельца, оболочки которых образованы фосфолипидами.
6) Метод биологической баллистики[6]. Это один из самых эффективных методов трансформации однодольных растений. Исходный материал для трансформации – суспензионная культура, каллусная ткань или 4–5-дневные культивируемые незрелые зародыши однодольных. Метод основан на напылении ДНК-вектора на мельчайшие частички вольфрама, которыми затем бомбардируют клетки. Бомбардировка осуществляется с помощью биолистической пушки за счет перепада давления. Часть клеток гибнет, а выжившие клетки трансформируются, затем их культивируют и используют для регенерации растений.
Решение проблемы создания новых форм растений подразумевает в первую очередь повышение качества синтезируемых растением продуктов, которые определяют его питательную и техническую ценность. В основном это касается запасных белков.
Начиная с 1970 г. стали появляться серьезные работы по изучению генов азотфиксации и их переносу в клетки Klebsiella pneumoniae и E.coli. Конструирование плазмид, несущих nif-гены, позволяет передавать способность к фиксации азота организмам, не обладающим этим свойством. Среди бактерий, кроме E.coli, такой перенос осуществлен для бактерий Salmonella typhimurium, Erwinia herbicola и других. Однако подобные манипуляции могут приводить к нежелательным эффектам. Так перенос генов в штамм Erwinia (бактерии, вызывающие гниение растений) может усилить его патогенное действие.
В настоящее время внимание ученых привлекают проблемы введения генов азотфиксации в клетки растений; создания ризоценозов между небобовыми растениями (особенно злаками) и азотфиксирующими организмами. Наиболее интересна первая проблема – введение nif-генов в клетки растений. Однако её решение сопряжено с рядом трудностей. Основная – разрушение нитрогеназы под воздействием кислорода. У азотфиксирующих микроорганизмов существует ряд приспособлений, защищающих бактерии от свободного кислорода. Таким образом, введение только nif-генов в какую-то растительную клетку не решает проблемы. Кроме того, сама клетка, в которую переносят гены азотфиксации, может быть не приспособлена к синтезу и расходованию большого количества энергии, которое требуется для фиксации азота. Следовательно, более перспективно повышение эффективности фиксации азота в уже существующих природных системах за счет воздействия на гены, контролирующие этот процесс, а также увеличение мощности корневой системы бобовых растений и создание новых азотфиксирующих систем с помощью методов клеточной инженерии.
Наибольший урон растениям приносят грибные, бактериальные и вирусные патогенны. В растении существуют защитные механизмы, которые в большей или меньшей степени (в зависимости от устойчивости растений) начинают действовать в ответ на проникновение фитопатогенов в клетку. Во-первых, начинается синтез соединений, вызывающих гибель патогенов. Примером могут служить специфические белки PRP (pathogen related proteins). Из них наиболее изучены ферменты хитиназы и β-1,3 – глюконазы, которые угнетают рост грибов и некоторых видов бактерий, разрушая их клеточные стенки. Во-вторых, могут создаваться структурные барьеры, препятствующие распространению инфекции. Это достигается благодаря лигнификации клеточных стенок.
Так, гены хитиназы и глюконазы кодируются одиночными генами. Благодаря этому были получены трансгенные растения табака и турнепса, в состав генома которых ввели ген хитиназы. Лабораторные и полевые испытания выявили большую устойчивость трансгенных растений. В растения томатов был введен ген защитных пептидов редьки (дефензинов) rs, отвечающих за устойчивость к фитопатогенным грибам.
Другой подход к получению трансгенных растений, устойчивых к вирусной инфекции, состоит во введении в геном исходных растений гена оболочки вируса. Это приводит к ингибированию размножения вируса и снижению инфицированности. Благодаря такому подходу был получен стойкий антивирусный эффект у растений табака, трансформированных геном оболочки вируса табачной мозаики (ВТМ).
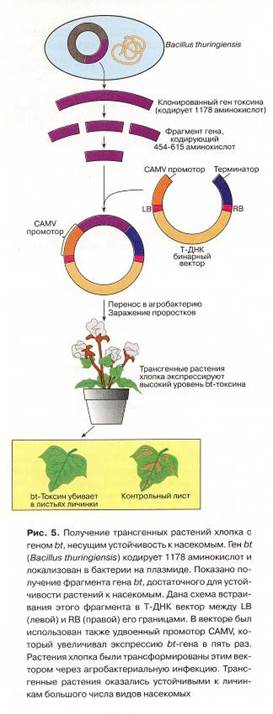
Создание трансгенных растений, устойчивых к насекомым, с помощью методов генной инженерии стало возможным после того, как было обнаружено, что бактерии Bacillus thurengiensis синтезируют специфический белок – прототоксин, высокотоксичный для насекомых. Попадая в кишечник насекомого, этот белок расщепляется, образуя активную форму токсина. В результате насекомое погибает. Ген, ответственный за экспрессию прототоксина, удалось обнаружить, выделить из генома B. thurengiensis и с помощью бинарного вектора ввести в геном растений табака.
Аналогичным образом растения томата были трансформированы генами другого инсектицидного белка – эндотоксина. В итоге были получены первые трансгенные растения, которые не повреждали насекомые (рис. 9).
Исследователи Колорадского университета (США) выяснили, что повреждению растений при замерзании способствуют бактерии эпифитной (поверхностной) микрофлоры Pseudomonas syringae и Erwinia herbicola, белки которых служат центрами кристаллизации[19]. Если обезвредить бактерии стрептомицином, то растения не замерзают при температуре -8ºС. Но стрептомицин дорог и вреден, поэтому выгоднее было изменить генетику данного штамма бактерий, вырезав из генома определенный ген. Растения, инфицированные мутантным штаммом P. syringae, росли при отрицательной температуре. Однако оказалось, что бактерии мутантного штамма более живучи и способны вытеснить природный штамм, который, попадая в верхние слои атмосферы, способствует кристаллизации атмосферной влаги. Вероятно, уничтожение природного штамма могло бы привести к экологической катастрофе.
Нельзя не сказать о «неопределённом» влиянии ГМ – продуктов на животные объекты. Всё же отмечены факты: у крыс, которых в течении девяти месяцев кормили ГМ – картофелем, произошло стойкое нарушение иммунной системы и ЖКТ.
Божьи коровки, которые питались тлями, жившими на ГМ – картофеле, становились бесплодными и т.д. Некоторые авторы связывают данные явления с «горизонтальными» потоками генетической информации[19].
Генную терапию на современном этапе можно определить как лечение наследственных, мультифакторных и ненаследственных (инфекционных) заболеваний путем введения генов в клетки пациентов с целью направленного изменения генных дефектов или придания клеткам новых функций[2]. Первые клинические испытания методов генной терапии были предприняты 22 мая 1989 года с целью генетического маркирования опухоль-инфильтрующих лимфоцитов в случае прогрессирующей меланомы. Первым моногенным наследственным заболеванием, в отношении которого были применены методы генной терапии, оказался наследственный иммунодефицит, обусловленный мутацией в гене аденозиндезаминазы (ADA). 14 сентября 1990 года в Бетесде (США) четырехлетней девочке, страдающей этим достаточно редким заболеванием (1: 100 000), были пересажены её собственные лимфоциты, предварительно трансформированные вне организма (ex vivo) геном ADA (ген ADA + ген neo + ретровирусный вектор). Лечебный эффект наблюдался в течение нескольких месяцев, после чего процедура была повторена с интервалом 3–5 месяцев. В результате лечения состояние пациентки настолько улучшилось, что она смогла вести нормальный образ жизни и не бояться случайных инфекций.