Лиственные мхи
Моховидные (Bryophyta).
Высшие споровые растения, представлены 25 тыс. видами. Произошли от водорослей, вероятно одновременно с риниофитами, предками папоротникообразных. Но это тупиковая ветвь, они никогда не имеют корней, воду поглощают выростами эпидермиса – ризоидами и у них нет настоящих проводящих тканей.
В жизненном цикле преобладает гаплоидный гаметофит, половое поколение. Это – единственная в истории растительного мира линия эволюции, связанная с регрессивным развитием спорофита! Спорофит (2n) (бесполое поколение) представлен спорангием на ножке, нижняя часть которой превращена в присоску, внедренную в тело гаметофита.
Из проросшей споры развивается протонема похожая на нитчатую водоросль, на ней развиваются почки, из которых вырастают листостебельные гаметофиты. На женском гаметофите образуется архегоний – женский половой орган, на мужском гаметофите – антеридии – мужские половые органы. В архегонии, который имеет бутыльчатую форму, образуется яйцеклетка, в антеридиях – сперматозоиды, которые по воде достигают яйцеклетки, из зиготы через несколько месяцев прорастает спорофит, который, например, у кукушкина льна, представляет собой коробочку на длинной ножке. В данном случае нижняя часть ножки превращена в гаусторий (присоску), внедряющийся в тело гаметофита. Спорофит лишен хлорофилла и полностью зависит от гаметофита ("паразитирует" на нем). Образованию спор предшествует редукционное деление – мейоз, при этом происходит редукция хромосом вдвое, и, следовательно, споры имеют гаплоидный набор хромосом. Затем спора прорастает и цикл замыкается. Мхи могут быть однодомными и двудомными. Однодомные – это те, у которых половые органы (антеридии и архегонии) расположены на одном растении, у двудомных – на разных.
Распространены повсюду, предпочитают увлажненные места. Небольшие растения, от 1 мм до 60 см. У листостебельных элементы проводящей ткани сходны с трахеидами и ситовидными трубками.
Моховидные объединяют в три класса:
Кл. антоцеротовые (Anthocerotopsida);
Кл. печеночники, или печеночные мхи (Marchantiopsida, или Hepaticopsida)
И самый многочисленный класс – листостебельные мхи (Bryopsida, или Musci), он, в свою очередь, делится на два подкласса:
Пор. Зеленые или настоящие мхи (Bryopsida) и
Пор. Белые, сфагновые или торфяные мхи (Sphagnida).
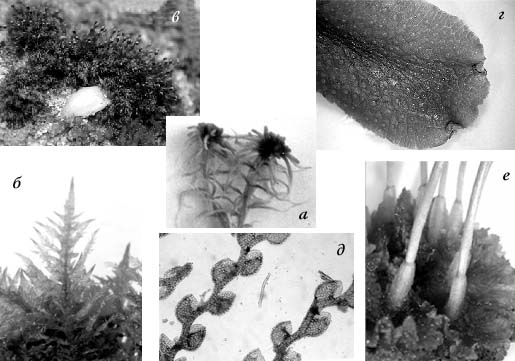
Рис 1. Представители основных групп мохообразных: а – Sphagnidae; б – Bryopsida; в – Andreidae; г – Jungermanniidae; д – Marchantiidae; е – Anthocerotopsida.

Рис 2. Формы роста мхов: а – наземные, б – эпифитные, в – водные, г – полуводные
Класс Листостебельные мхи, или Бриопсиды (Bryopsida, или Musci).
Лиственные мхи – весьма обширная группа, насчитывающая в своем составе около 13 тысяч видов, распространенных по всему свету и живущих в самых разнообразных условиях. По величине и своему внешнему виду они очень разнообразны, но притом они всегда сохраняют свой характерный облик, по которому их очень легко отличить от всякого другого растения. Все они имеют округлый стебель, густо покрытый внешними листьями, сидящими тесной спиралью и собранными на вершине их в виде почки. Редко стебель остается простым, большей частью он ветвится, причем ветвление иногда достигает большой сложности. Оно может быть и моноподиальным и симподиальным. В почве стебель укореняется при помощи ризоидов, представляющих собой длинные, обильно разветвленные однорядные нити; иногда они, как у Polytrichum, скручиваются в жгуты. Размеры мхов колеблются в широких пределах; наиболее мелкие из них измеряются миллиметрами, самые крупные (Dawsonia из семейства Polytrichaceae) достигают 0,5 м в высоту. При всем внешнем разнообразии лиственных мхов внутреннее строение их остается простым. Настоящих сосудистых пучков у них никогда не образуется. Кукушкин лен является одним из наиболее сложно устроенных мхов, и его проводящий пучок показывает высшую степень сложности, какой достигает среди мхов проводящая система. У громадного большинства мхов проводящий пучок, занимающий центр стебля, состоит из очень узких, вытянутых в длину, тонкостенных клеточек. Остальная часть стебля (кора) построена из паренхиматических клеток, принимающих иногда ближе к периферии характер клеток механических. С поверхности стебель покрыт эпидермисом. У очень многих мхов проводящего пучка совершенно не образуется, и весь стебель состоит только из паренхимы. Листья у громадного большинства мхов являются однослойными пластинками, лишь в средней жилке клетке располагаются в несколько слоев.
Среди лиственных мхов весьма развита способность к вегетативному размножению. Оно совершается или при помощи подземных побегов и плетей, когда концы их вырастают в новые особи, или при помощи специальных выводковых почек. Эти последние образуются на стеблях, листьях или же в особых вместилищах, возникающих на стеблях. Выводковые почки имеют самую разнообразную форму.
Попадая в благоприятные условия, они прорастают в протонему, на которой уже возникают облиственные стебли. Для вегетативного размножения может служить и сама протонема, когда на ней образуется большое количество почек. Половыми органами у Bryopsida являются антеридии ♂ и архегонии ♀. В большинстве случаев они располагаются отдельно друг от друга на вершине стеблей, образуя мужские и женские «цветки». Бесполым поколением является спорогоний. Он всегда состоит из ножки и коробочки и покрыт сверху колпачком, но все эти составные части у различных мхов весьма варьируют и по величине и по форме. Длина ножки, величина и форма колпачка, положение спорогония на стебле – все это признаки, имеющие важное систематическое значение при классификации мхов. Весьма важную роль в классификации их играет и перистом, т.е. те зубцы, которые окружают отверстие урны, после того как спадет крышечка; при этом во внимание принимаются расположение, форма и строение его зубцов.
Главными подклассами класса являются:
1) Bryopsida – настоящие мхи
2) Sphagnida – сфагновые, белые или торфяные мхи.
Порядок сфагновые, белые или торфяные мхи (Sphagnida.)
|
Рис 3. Растения сфагнов, объединенные в дернинку |
Рис 4. Головки сфагновых мхов |


Морфология
Стебель у сфагнов прямостоячий, но отдельные растения не способны расти прямо из-за отсутствия механической ткани. Поэтому сфагнум всегда растет дернинками (рис 4), в которых отдельные растения опираются друг на друга веточками, растущими из стебля горизонтально, подобно тому, как могут опираться, обнявшись, друг другу на плечи люди. На верхушке стебля находится более или менее компактная головка, состоящая из скученных вокруг точки роста коротких молодых веточек (рис 3). По мере роста стебля вверх веточки вырастают в длину и рассредоточиваются по стеблю отдельными пучками, состоящими из горизонтально отстоящих и свисающих вдоль стебля веточек (рис 5). Отстоящими веточками соседние растения в дернинке сцепляются между собой. По пористым клеткам листьев этих веточек осуществляется горизонтальный транспорт веществ в сфагновой дернинке. По клеткам листьев свисающих ветвей осуществляется транспорт веществ по вертикали (из почвенного раствора). Листостебельное растение сфагна не имеет ризоидов.