Популяционно-генетическая структура вида у гидробионтов
Содержание
Владивосток
1. Основные внутривидовые группировки животных
ОБЩИЙ ГЕНЕТИЧЕСКИЙ ПОДХОД: ПЕРСПЕКТИВЫ И ОГРАНИЧЕНИЯ
Перспективы и ограничения генетического подхода
Средняя гетерозиготность
Генетическое расстояние. Стандартное расстояние Нея (Dn)
Определение вида
2. Специфика популяционно-генетической структуры у рыб и беспозвоночных. Факторы популяционно-генентической динамики и стационарности. Генетические основы рационального использования природных ресурсов
Феноменологический подход
Научный подход
Главные факторы популяционно-генетической динамики
Мера межпопуляционной дифференциации
Простейшая модель популяционной структуры
Комплексная природа генетической динамики и стационарности
Классификация вариантов внутривидовой структуры, на основе использования популяционно-генетических средств
Особенности популяционной организации морских организмов
Генетическая изменчивость и дифференциация
ПРИМЕР ПОПУЛЯЦИОННО-ГЕНЕТИЧЕСКОГО ИССЛЕДОВАНИЯ
3. Модели популяционно-генетической организации вида
Модель панмиктической популяции
Модели подразделенной популяции
Островная модель
Модель ступенчатой миграции
ЭМПИРИЧЕСКИЕ ОЦЕНКИ ПАРАМЕТРОВ 20
1. Основные внутривидовые группировки животных
ОБЩИЙ ГЕНЕТИЧЕСКИЙ ПОДХОД: ПЕРСПЕКТИВЫ И ОГРАНИЧЕНИЯ
Проблемы биологического вида и видообразования привлекают исследовательский интерес с момента становления биологии, как науки. Данные проблемы являются одними из ключевых в общей биологии и общей генетике. Наиболее принята в настоящее время синтетическая теория эволюции (СТЭ), частью которой является биологическая концепция вида (БКВ).
Перспективы и ограничения генетического подхода
Перспективы.
Редукция данных до генотипических значений (кодов) дает возможность использовать генетическую теорию в анализе.
Возможно сравнительное исследование изменчивости между таксонами по структурным и регуляторным элементам генома.
Исследование дивергенции видов из природы по белкам и ДНК привело к открытию «молекулярных часов».
Появилась возможность филогенетических реконструкций: (1) не по сходству, а (2) по родству, за счет оценки хронологии и последовательности ветвлений в молекулярных филетических линиях.
Ограничения.
Дедукция ограничена генотипическими описаниями и генетической теорией.
Анализ связан с предварительным, трудоемким экспериментальным исследованием (со своими собственными ограничениями).
Исследование видов из природы часто ограниченно уникальностью или редкой повторяемостью события (феномена).
Генотипические эффекты маркерных локусов на фенотип являются слабыми.
Теория по некоторым направлениям не достаточно развита.
В целом необходимы данные такого рода:
Данные, которые подтверждают (отвергают) центральную догму неодарвинизма – эволюция может происходить только на основе генетических изменений.
Данные об изменчивости на различных уровнях биологической организации в генетических терминах (полокусные генотипические значения – AA, …) – одномерные таблицы данных (ТД).
Данные о генотипических значениях для особей по набору локусов (комплексный генотип – AA Bb…) – многомерные ТД.
Дополнительные информация: морфологические данные, данные об абиотической изменчивости (по меньшей мере, как экспертные оценки – группирующие переменные) и др.
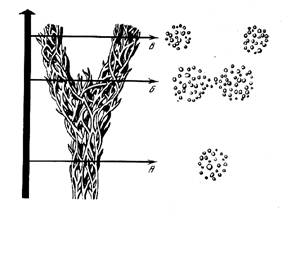 |
Рис. 0. Схематическое представление видовой дивергенции во времени. А – единая видовая популяция, B – начальная фаза дивергенции (подвиды), C – различные виды. Кружки справой стороны представляют объединения отдельных демов или популяций, между которыми имеется поток генов вплоть до фазы В (По Dobzhansky, 1955). Наиболее представительными в настоящее время являются данные по изменчивости белковых или точнее ферментных генов. |
Посредством генетико-биохимического подхода и некоторых популяционно-генетических формул легко вычислить как уровень генетической изменчивости, так и степень сходства (или различия) генофондов особей из популяции одного вида или из различных видов.
Средняя гетерозиготность.
H = Σ Li=1 hk / L.
H – это средняя ожидаемая гетерозиготность на локус или на особь.
hk = 1 - Σ mi=1 pi2, hk – ожидаемая гетерозиготность по одному локусу (Hexp=Hs), pi – частота i–того аллеля; L – число локусов. Наблюдаемая гетерозиготность равна: Hobs = hj / n, hj – число гетерозигот в выборке, n – общее число особей в выборке.
Генетическое расстояние. Стандартное расстояние Нея (Dn).
Dn = - ln I.
I – нормализованная генетическая идентичность для случайной выборки локусов. Для локуса j она равна:
Ij = (Σ ki=1 xi yi) / √ Σ ki=1 xi 2 Σ ki=1 yi 2
Для всего набора локусов I равно:
I = Jxy / √ JxJy;
где Jxy, Jx, Jy – средние арифметические для сумм: Σ ki=1 xi yi, Σ ki=1 xi2, Σ ki=1 yi2.
Здесь, xi и yi – это частоты аллелей в популяциях (таксонах) x и y, k – число аллелей во всех попарных сравнениях.
Определение вида.
Вид – это биологическая группировка, состоящая из одной или нескольких популяций перекрестно размножающихся особей, которые репродуктивно изолированы от других таких группировок и стабильны в природе, занимая определенный ареал. В принципиальных позициях, это определение, типичное для БКВ. Так, одно из первоначальных определений БКВ формулируется следующим образом “Вид – это репродуктивное сообщество популяций (репродуктивно изолированных от других), которое занимает определенную нишу в природе” (Mayr, 1982, p. 273). БКВ наиболее близко подходит к теории популяционной генетики. Имеются несколько других концепций вида со своими преимуществами и ограничениями, которые были критически проанализированы (Красилов, 1977; King, 1993). Список этих концепций дан ниже.
- Линнеевский вид.
- Биологическая концепция вида.
- Биологическая концепция вида в модификации Майра (БКВ) (Майр, 1947; 1968)
- БКВ, модификация II (Mayr, 1982).
- Концепция «распознания» (recognition ) вида (Paterson, 1978; 1985).
- Концепция сплоченности (cohesion) вида (Templeton, 1989).
- Эволюционная концепция вида.
- Эволюционная концепция вида Симпсона (Simpson, 1961).
- Эволюционная концепция вида Вилея (Wiley, 1978).
- Экологическая концепция вида (Van Valen, 1976).
- Филогенетическая концепция вида (Cracraft, 1983).