Наследственность, представления о генетическом коде, гены индивидуальности
Развитие цитологии пртивело к постановке вопроса о материальных основах наследственности. Впервые мысль о роли ядра как носителя наследственности была сформулирована
О. Гертвигом (1884) и Э. Страсбургером(1884) на основании изучения процесса оплодотворения. Т. Бовери (1887) установил индивидуальность хромосом и развил гипоьезу о их качественном различии. Он же, а также Э. ван Бенедет (1883) установили уменьшение количества хромосом вдвое при образовании половых клеток в мейозе. Американский ученый У. Сеттон (1902) дал цитологическое объяснение закону Менделя о независимом наследовании приизнаков. Однако подлинное обоснование хромосомной теории наследственности было дано в работах Т. Моргана и его школы (начиная с 1911), в которых было показано точное соответствие между генетическими и цитологическими данными. В опытах на дрозофилебыло установлено нарушениет независимого распределения признаков-их сцепленное наследование. Это явление было объяснено сцеплением генов, то есть нахождением генов, определяющих эти признаки, в одной определенной паре хромосом. Изучение частоты рекомбинаций между сцеплеными генами (в результате кроссинговера) позволило составить карты расположения генов в хромосомах.
Генетические карты хромосом - схемы относительного расположения сцепленных между собой наследств. факторов — генов. Генетические карты хромосом отображают реально существующий линейный порядок размещения генов в хромосомах и важны как в теоретических исследованиях, так и при проведении селекционной работы, т. к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. Имея Генетические карты хромосом, можно по наследованию «сигнального» гена, тесно сцепленного с изучаемым, контролировать. передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; напр., ген, определяющий сморщенный эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения. Многочисленные факты отсутствия (вопреки законам Менделя) независимого распределения призна
ков у гибридов второго поколения были объяснены хромосомной теорией наследственности. Гены, расположенные в одной хромосоме, в большинстве случаев наследуются совместно и образуют одну группу сцепления, количество к-рых, таким образом, соответствует у каждого организма гаплоидному числу хромосом. Американский генетик Т. X. Морган показал, однако, что сцепление генов, расположенных в одной хромосоме, у диплоидных организмов не
абсолютное; в нек-рых случаях перед образованием половых клеток между однотипными, или гомологичными, хромосомами происходит обмен соответств. участками; этот процесс носит назв. перекреста, или кроссинговера. Обмен участками хромосом (с находящимися в них генами) происходит с различной вероятностью, зависящей от расстояния между ними (чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации). Генетич. анализ позволяет обнаружить перекрест только при различии гомологичных хромосом по составу генов, что при кроссинговере приводит к появлению новых генных комбинаций. Обычно расстояние между генами на Генетических картах хромосом выражают как процент кроссинговера (отношение числа мутантных особей, отличающихся от родителей иным сочетанием генов, к общему кол-ву изученных особей); единица этого расстояния — морганида — соответствует частоте кроссинговера в 1 %.
Итак, выделим основные положения хромосомной теории наследственности:
1. Гены располагаются в хромосомах, различные хромосомы содержат неодинаковое чис ло генов, набор генов каждой из негомологичных хромосом уникален.
2. Гены в хромосоме расположены линейно, каждый ген занимает в хромосоме определенный локус (место).
3. Гены , расположенные в одной хромосоме, образуют группу сцепления и вместе (сцеплено) передаются потомкам, число групп сцепления равно гаплоидному набору хромосом.
4. Сцепление не абсолютно, так как в профазе мейоза может происходить кроссинговер и гены, находящиесяв одной хромосоме, разобщаются. Сила сцепления зависит от расстояния между генами в хромосоме: чем больше расстояние, тем меньше сила сцепления. и наоборот. Расстояние между генами измеряется в процентах кроссинговера. 1% кроссинговера соответтствует одной морганиде.[4]
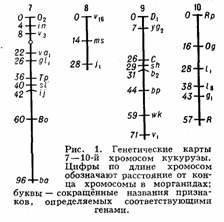
Генетические карты хромосом составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые назв. мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры. Составить Генетические карты хромосом можно только для объектов, у которых изучено большое число мутантных генов. Например, у дрозофилы идентифицировано свыше 500 генов, локализованных в её 4 группах сцепления, у кукурузы — около 400 генов, распределённых в 10 группах сцепления (рис. 1). У менее изученных объектов число обнаруженных групп сцепления
меньше гаплоидного числа хромосом. Так, у домовой мыши выявлено около 200 генов, образующих 15 групп сцепления (на самом деле их 20); у кур изучено пока всего 8 из 39. У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом.
|
|
|
|
|
|
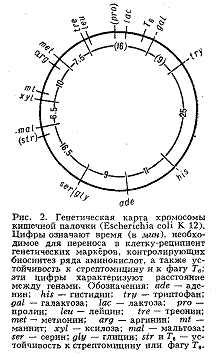
У бактерий, к-рые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления (рис. 2). При переносе генетич. материала из клетки-донора в клетку-реципиент, например при конъюгации, кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110-120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбинантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Генетических карт хромосом бактерий, детально разработанных у ряда видов. Ещё более детализированы Генетические карты хромосом нек-рых бактериофагов
Генетика пола. Количество групп сцепленных генов оказалось равным количеству пар хромосом, присущих данному виду. Важнейшие доказательства хромосомной теории наследственности были получены при изучении наследования, сцепленного с полом. Ранее цитологи открыли в хромосомных наборах ряда видов жиивотных особые , так называемые половые хромосомы, которыми самки отличаются от самцов. В одних случаях самки имеют 2 одинаковые половые хромосомы(XX), а самцы-разные(XY), в других - самцы-2 одинаковые(XX, или ZZ), а самки - разные(XY, или ZW). Пол с одинаковыми половыми хромосомами называется гомогаметным , с разными - гетерогаметным. Женский пол гомогаметен , а мужской гетерогаметен у некоторых насекомых ( в том числе у дрозофилы) и всех млекопитающих. Обратное соотношение - у птиц и бабочек. Ряд признаков у дрозофилы наследуется в