Атф индуцированное изменение внутриклеточной концентрации кальция в нейронах неокортекса крыс
IP3-управляемые Са2+ каналы ЭР. Существование IP3-управляемых Са2+ каналов впервые было обнаружено в нейронах Пуркинье. Позже было показано, что они встроены в мембрану эндоплазматического ретикулума. Структура IP3-управляемых Са2+ каналов сходна со структурой Са2+-управляемых Са2+ каналов ЭР. Они также являются гомотетрамерами с молекулярным весом мономера 260 КД. 50% этих каналов активируется 15 мкмоль/л IP3 и блокируется рутением красным и La3+. IP3-управляемые Са2+ каналы были выделены из мозга млекопитающих, и их аминокислотная последовательность была расшифрована. Было показано, что семейство генов, экспрессирующих IP3-управляемые Са2+ каналы, состоит из трех или четырех различных генов; они характеризуются различной чувствительностью к IP3 и по-разному распределены в мозге млекопитающих (45). Порог активации этих каналов варьирует между 0.2 - 0.5 мкмоль/л в нейронах Пуркинье мозжечка и возрастает до 9 мкмоль/л в астроцитах.
2.1.4 Кальциевые насосы
Существует два семейства Са2+ насосов, ответственных за устранение ионов Са2+ из цитоплазмы: Са2+ насосы плазмалеммы и Са2+ насосы эндоплазматического ретикулума. Хотя они относятся к одному семейству белков (так называемому P-классу АТФ-аз), эти насосы обнаруживают некоторые различия в строении, функциональной активности и фармакологии.
Кальциевый насос плазмалеммы. Са2+ насос плазмалеммы, который удаляет ионы Са2+ из цитоплазмы в межклеточное пространство, был открыт в 1966 году. Молекулярные свойства Са2+ насосов плазмалеммы описаны в нескольких обзорах (18), однако достоверных данных о скорости вывода Са2+ и регуляции Са2+ насосов в нервных клетках немного. Недавно был разработан двухфлуоресцентный микрокапельный метод (58), позволяющий одновременно измерять [Ca2+]i и выход Са2+ наружу на одиночных клетках. Исследования, проведенные с помощью данного метода на нейронах моллюска и секреторных клетках, показали, что активность Са2+ насоса плазмалеммы контролируется непосредственно [Ca2+]i: увеличение концентрации цитоплазматического кальция активирует Са2+ насос (58). В нейронах моллюска около 40% ионов кальция, входящих в клетку в ответ на деполяризацию мембраны, выводится из нейрона уже во время фазы нарастания [Ca2+]i, отражая таким образом активацию кальциевого насоса плазмалеммы увеличением концентрации цитозольного Са2+ (58).
Кальциевый насос эндоплазматического ретикулума. Во многих эукариотических клетках, наряду с Са2+ насосом плазмалеммы, существует кальциевый насос сарко(эндо)плазматического ретикулума (SERCA). В настоящее время описано по крайней мере 3 различных изоформы SERCA-насосов в клетках млекопитающих. SERCA1-подтип сосредоточен исключительно в быстрых скелетных мышцах, SERCA2-насосы широко распространены в других тканях. Значимость SERCA3-насосов менее ясна (13). Белки SERCA2-насосов разделяются на две различные изоформы: SERCA2а, характерные для кардиомиоцитов и гладких мышц, и SERCA2b, характерные для тканей мозга. Предполагается, что насосы SERCA различными способами регулируются цитоплазматической и интралюминальной концентрациями Са2+: Увеличение [Ca2+]i активирует захват ионов кальция в ЭР, в то время как увеличение свободного кальция внутри ЭР ингибирует насосы SERCA (12). Насосы SERCA эффективно и селективно блокируются тапсигаргином в наномолярных концентрациях (37) и микромолярными концентрациями циклопиазоновой кислоты. Однако, тапсигаргин вызывает также блокаду потенциал - управляемых кальциевых каналов плазмалеммы, как это показано на клетках коркового слоя надпочечников и на сенсорных нейронах (Shmigol et al., 1995), поэтому его следует использовать с некоторой осторожностью.
2.1.5 Кальциевые обменники
Дополнительным механизмом, ответственным за вывод ионов кальция из цитоплазмы, является натрий-кальциевый обменник, который выводит Са2+, используя энергию натриевого электрохимического градиента. Наличие Na+- Са2+ обменника было показано в различных типах возбудимых и невозбудимых клеток; в клетках нервной системы он был обнаружен в конце 60-х годов (9). В нейронах моллюска, помещенных в среду с пониженным натрием (т.е. с обратным натриевым градиентом), наблюдалось увеличение [Ca2+]i, что является результатом работы обменника в инвертированной форме. Однако, вклад Na+- Са2+ обменника в регуляцию [Ca2+]i в нейронах млекопитающих до сих пор не оценен. В некоторых работах было показано, что обменник принимает незначительное участие в удалении цитоплазматического Са2+, в то время как в других работах представлены данные о том, что обменник играет существенную роль в переносе Са2+ через мембрану (57).
2.1.6 Са2+-связывающие органеллы
Кроме быстрого связывания цитозольного Са2+ внутриклеточными Са2+-связывающими белками, ионы кальция, попадающие в цитозоль, могут аккумулироваться аппаратом Гольджи или клеточным ядром, захватываться митохондриальными Са2+ депо, имеющими достаточно невысокое сродство к Са2+, или быстрыми депо, связанными с ЭР или СР, имеющими высокое сродство к Са2+. Однако если [Ca2+]i превышает 0,5 мкмоль/л, наблюдается существенное перераспределение [Ca2+]i в область митохондрий. Буферные системы митохондрий принимают участие в удалении избыточного Са2+ из цитоплазмы в клетках кишечника, некоторых типах нервных клеток (59) и в секреторных клетках после повышения [Ca2+]i, стимулированного агонистами. Связывание кальция митохондриями обеспечивается активностью систем, расположенных на внутренней митохондриальной мембране. Са2+ поступает в митохондрии по электрохимическому градиенту; разность потенциалов, обеспечивающая транспорт кальция, создается переносом электронов во время клеточного дыхания и связанного с ним переносом протонов. Перенос электронов по дыхательной цепи является основным механизмом, обеспечивающим энергетику транспорта кальция. Подавление дыхательной цепи карбонил-цианид-м-хлорофенил-гидразоном (СССР) эффективно блокирует аккумуляцию кальция митохондриями (41).
2.2 Влияние АТФ на кальциевый гомеостаз
Последние исследования показали, что АТФ занимает прочное место в ряду нейромедиаторов центральной и периферической нервной систем (Burnstock 1990). Не вызывает сомнения, что АТФ является не только важнейшим внутриклеточным метаболитом, но и служит важным объектом межклеточного взаимодействия.
2.2.1 Строение и свойства АТФ
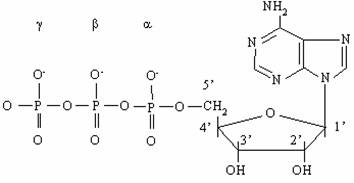
|
АТФ (см. рис.1) представляет собой нуклеотид и как всякий нуклеотид состоит из трех компонентов: азотистого основания, сахара пентозы и фосфата. В качестве азотистого основания в нуклеотидах присутствуют производные пурина и пиримидина. Фосфаты соединены в полифосфатную цепь, количество которых в естественных нуклеотидах не превышает трех. Однако синтезированы нуклеотиды, содержащие линейные цепи из более чем 3-х фосфатов, к примеру аденозинтетра- и аденозинпентафосфаты.