Атф индуцированное изменение внутриклеточной концентрации кальция в нейронах неокортекса крыс
С целью обеспечения достоверности полученных данных, принимая во внимание тот факт, что работа проводилась на тонких срезах мозга, мы провели серию экспериментов с подавлением распостранения потенциалов действия с помощью тетродотоксина (TTX). При использовании TTX выяснилось, что амплитуда и форма [Ca2+]i транзиентов не изменяется или изменяется незначительно. Полученные результаты позволяют нам предполагать, что получаемые нами данные отображают процессы происходящие в отдельной наблюдаемой клетке. К сожалению мы не имели возможности проводить все эксперименты c тетродотоксином из-за дороговизны препарата. Для изучения характеристик рассматриваемых пуринорецепторов, принимающих участие в генерации [Ca2+]i транзиента, применялись следующие протоколы экспериментов.
1. Приложение АТФ в растворе не содержащем Ca2+. Удаление кальция из внеклеточного раствора незначительно влияло на кальциевый транзиент, вызванный аппликацией 10 мкМ АТФ, но в тоже время подавляло на 45% ± 7% кальциевый транзиент вызванный 100 мкМ АТФ. (Рис. 8).
|
|
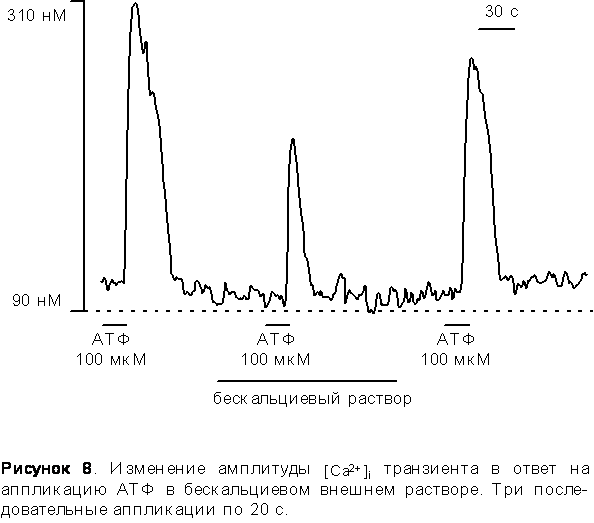
1. Для исследования способности внутриклеточного Ca2+ депо к перезаполнению мы последовательно апплицировали АТФ несколько раз с интервалом в 60 секунд в обычном растворе и в растворе не содержащем Ca2+ . Как видно из рисунка 9 вторая аппликация АТФ вызывала Ca2+ транзиент меньшей (на 30% ± 4%) амплитуды по сравнению с первой (контрольной) аппликацией. Последующие аппликации АТФ разделенные 60 ти секундным интервалом не вызывали заметного уменьшения амплитуды [Ca2+]i транзинета по сравнению со второй аппликацией в стандартном растворе
|
|

В бескальциевом растворе - наоборот, вторая аппликация АТФ вызывала значительное уменьшение Ca2+ транзиента на 40% ± 5% от контроля. Последующие аппликации АТФ в бескальциевом растворе приводили к последовательному уменьшению амплитуды [Ca2+]i транзиента и после двух - трех последовательных аппликаций сигнал исчезал вообще (рисунок 10). Такое уменьшение вероятно обусловлено истощением внутриклеточных депо.
|
|

1. Ингибирование захвата Ca2+ эндоплазматическим ретикулом тапсигаргином (спецефическим блокатором Ca2+ насоса эндоплазматического ретикулума) уменьшало [Ca2+]i транзиенты, вызванные приложением АТФ, на 63% ± 5% для концентрации АТФ 100 мкМ АТФ (рис. 11). Как видно из рисунка 11 приложение тапсигаргина не влияло на базальный уровень Ca2+ в клетке.
1. Приложение кадмия в концентрации 50 мкМ, верапамила в концентрации 100 мкМ и 50 мкМ никеля обратимо уменьшало амплитуду [Ca2+]i транзиента вызванного приложением 100 мкМ АТФ на 35%, 20%, и 15% сответственно. Данные представлены на рисунке 12.
|
|
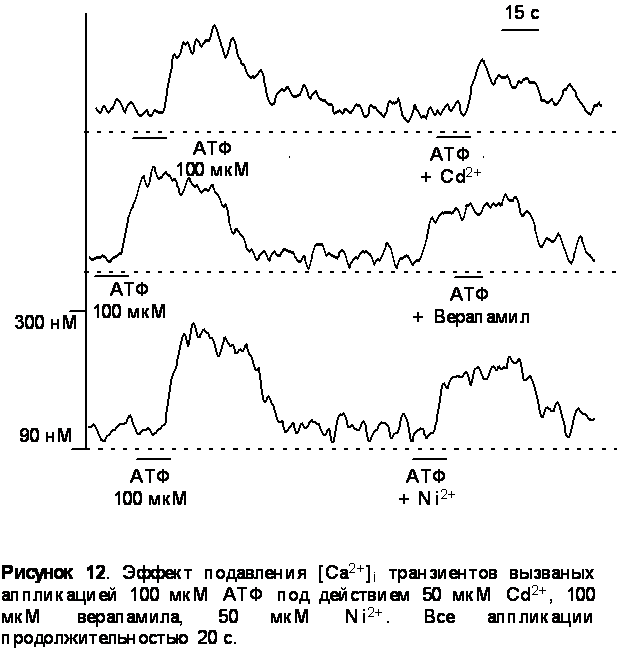
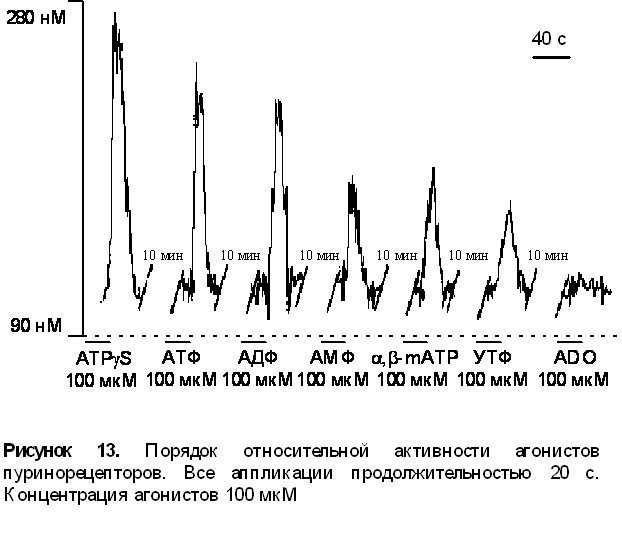
1. Приложение различных агонистов пуринорецепторов вызывало различные по амплитуде ответы. Порядок относительной активности лигандов для данного объекта был следующим ATPgS > ATФ = AДФ >> a,b-methylene ATP » AMФ > УТФ>> аденозин (ADO), данные преставлены на рисунке 13.
|
|
1. Сурамин, известный антагонист Р2 типа пуринорецепторов, уменьшал [Ca2+]i транзиенты, вызванные приложением 100 мкМ АТФ на 76% ± 7% и также не изменял концентрацию [Ca2+]i в покое. Данные представлены на рисунке 14.
|
|

5. ОБСУЖДЕНИЕ
При исследовании средних слоев неокортекса крыс мы обнаружили их способность отвечать на приложение АТФ. Таким образом мы можем сделать вывод о наличии в данном объекте пуринорецепторов. Ответ на АТФ является доза - зависимым с амплитудой половинного ответа в 220 нМ. Последовательные приложения АТФ, после второго приложения, не вызывали уменьшения амплитуды [Ca2+]in транзиентов. Из этого можно сделать вывод об отсутствии десенситизации данного типа рецепторов.
Исследуя источники повышения цитозольного кальция мы обнаружили, что АТФ активирует как ионотропные так и метаботропные рецепторы. Блокаторы потенциал - управляемых кальциевых каналов такие как кадмий, никель и верапамил уменьшали АТФ - индуцированные кальциевые транзиенты на 35% - 15%, что говорит об опосредованном АТФ активировании потенциал - управляемых каналов.
Для исследования активацию метаботропных рецепторов. Для этого мы прилагали АТФ в бескальциевом растворе, - амплитуда ответа при этом уменьшалась на 45% ± 7%. Этот результат говорит о том, что внутриклеточные депо принимают участие в генерации [Ca2+]in ответов, причем их вклад близок к половине. При повторных аппликациях АТФ в бескальциевом растворе Ca2+ ответ исчезал полностью после второй аппликации, т.е. внутриклеточные депо полностью истощаются. Исследуя путь высвобождения внутриклеточного кальция мы апплицировали тапсигаргин - спецефический блокатор АТФ - азы эндоплазматического ретикулума. [Ca2+]in транзиенты уменьшались на 63% ± 5% в присутствии тапсигаргина. Аппликация коффеина, агониста рианодиновых рецепторов, в клетках моторной коры 14 дневных крыс не вызывали повышения уровня [Ca2+]in . Следовательно выброс [Ca2+]in из внутриклеточного депо происходит по IP3 - чувствительному механизму.
В дальнейшем мы исследовали более подробно типы присутствующих пуринорецепторов. Построив ряд активности агонистов для Р1 и Р2 пуринорецепторов по амплитудам ответов, мы сделали заключение базирующееся на отсутствии ответа на аденозин, что в нашем объекте присутствуют только Р2 пуринорецепторы. Проводя дальнейшую субклассификацию Р2 типа рецепторов мы использовали сурамин - блокатор некоторых типов Р2х и Р2у рецепторов. Приложение сурамина уменьшало амплитуду [Ca2+]i транзинета вызванного приложением 100 мкМ АТФ на 76% ± 5%, что говорит о наличии этих типов рецепторов в исследуемом объекте.
6. ВЫВОДЫ
1. Приложение АТФ в различных концентрациях вызывает Ca2+ транзиенты в клетках моторной коры крыс.
2. Ответ на АТФ является доза - зависимым с амплитудой половинного ответа в 220 нМ.