Математические модели естествознанияРефераты >> Естествознание >> Математические модели естествознания
Сделаем ряд замечаний. При анализе уравнения (2) мы предполагали, что коэффициенты ![]() и
и ![]() не зависят от
не зависят от ![]() . Действительно для мембраны емкость практически постоянна. Однако, проводимость
. Действительно для мембраны емкость практически постоянна. Однако, проводимость ![]() есть функция от
есть функция от ![]() . Проведенный анализ справедлив лишь при малых отклонениях
. Проведенный анализ справедлив лишь при малых отклонениях ![]() от равновесного значения
от равновесного значения ![]() .
.
Пусть теперь дополнительно к ![]() в полости и в сосуде присутствует
в полости и в сосуде присутствует ![]() , концентрации которого в полости и в сосуде различны. Ионам
, концентрации которого в полости и в сосуде различны. Ионам ![]() отвечает свое равновесное значение
отвечает свое равновесное значение ![]() . Натриевый ток
. Натриевый ток ![]() связан с мембранным потенциалом
связан с мембранным потенциалом ![]() соотношением:
соотношением: ![]() . Привлекая снова закон Кирхгофа, получаем:
. Привлекая снова закон Кирхгофа, получаем:
![]()
![]() . . Cu¢ + gNa(u - ENa) +gK(u -EK ) (3)
. . Cu¢ + gNa(u - ENa) +gK(u -EK ) (3)
Теперь в силу (3) равновесное значение мембранного потенциала суть:
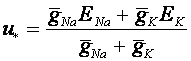 .
.
Одна из первых гипотез о природе мембранного потенциала была высказана в 1902 г. Бернштейном, который использовал положения электрохимии. Он предположил, что предположил, что концентрация ионов ![]() в клетке выше, чем во внеклеточной среде. Вследствие этого ионы
в клетке выше, чем во внеклеточной среде. Вследствие этого ионы ![]() диффундируют из клетки. Образуется их относительный дефицит положительных ионов внутри клетки и внутренняя поверхность мембраны заряжается отрицательно. Тем самым, потенциал покоя определяется равновесным электрохимическим потенциалом для ионов
диффундируют из клетки. Образуется их относительный дефицит положительных ионов внутри клетки и внутренняя поверхность мембраны заряжается отрицательно. Тем самым, потенциал покоя определяется равновесным электрохимическим потенциалом для ионов ![]() . Далее, Бернштейн предположил, что в жизни нейрона иногда наступает особый момент -состояние возбуждения, когда на короткое время мембрана становится проницаемой и для других ионов, в частности, для натрия и хлора. В результате притока внутрь клетки положительных ионов величина мембранного потенциала падает, а затем, после восстановления избирательных свойств мембраны, снова входит в норму. Так впервые была объяснена генерация спайка.
. Далее, Бернштейн предположил, что в жизни нейрона иногда наступает особый момент -состояние возбуждения, когда на короткое время мембрана становится проницаемой и для других ионов, в частности, для натрия и хлора. В результате притока внутрь клетки положительных ионов величина мембранного потенциала падает, а затем, после восстановления избирательных свойств мембраны, снова входит в норму. Так впервые была объяснена генерация спайка.
Натриево - калиевый цикл
Гипотеза Бернштейна хорошо объясняла многие известные физиологам того времени факты. В частности, зависимость потенциала покоя от концентрации калия в межклеточной среде, а также от температуры. Однако в то время гипотезу нельзя было подвергнуть экспериментальной проверке. В распоряжении исследователей не было методики непосредственного изменения мембранного потенциала и концентраций ионов. Гипотеза Бернштейна вызывала массу споров.
Ситуация изменилась после сообщения английского зоолога Дж. Юнга, сделанного в 1936 г. Он обнаружил, что длинные тяжи у кальмаров и каракатиц являются не кровеносными сосудами, как это считалось ранее, а необычайно толстыми аксонами (отростками нервных клеток). Они получили название гигантских аксонов и стали естественным объектом для изучения мембран. Диаметр аксонов достигает 1 мм и дает возможность вводить в них электроды и капилляры. В результате можно измерить как мембранный потенциал, так и концентрацию внутриклеточного раствора.
Выяснилось, что мембранная теория Бернштейна в целом верна, хотя и нуждается во внесении ряда существенных дополнений и изменений. В частности верным оказалось утверждение, что потенциал покоя обусловлен в основном электрохимическим потенциалом ионов ![]() . Наоборот, согласно теории Бернштейна в точке максимума потенциала действия (спайка) мембранный потенциал должен быть равен нулю (ток ионов прекращается, когда напряжение равно нулю). Однако, английские физиологи А. Ходжкин и А. Хаксли в 1939 г. показали, что на максимуме потенциала действия мембранный потенциал не уничтожается, а меняет знак. Внешняя поверхность мембраны заряжается положительно по отношению к внутренней. Явление получило название «овершут» и не имело объяснений в рамках теории Бернштейна. Оно связано с неравномерным распределением ионов натрия. Оказалось, что концентрация ионов
. Наоборот, согласно теории Бернштейна в точке максимума потенциала действия (спайка) мембранный потенциал должен быть равен нулю (ток ионов прекращается, когда напряжение равно нулю). Однако, английские физиологи А. Ходжкин и А. Хаксли в 1939 г. показали, что на максимуме потенциала действия мембранный потенциал не уничтожается, а меняет знак. Внешняя поверхность мембраны заряжается положительно по отношению к внутренней. Явление получило название «овершут» и не имело объяснений в рамках теории Бернштейна. Оно связано с неравномерным распределением ионов натрия. Оказалось, что концентрация ионов ![]() внутри клетки примерно в десять раз ниже чем в межклеточной жидкости. В начале спайка ионы натрия под действием электрического поля и концентрационного градиента устремляются из межклеточной среды в клетку (в клетке их концентрация ниже, а внутренняя поверхность мембраны заряжена отрицательно по отношению к внешней). В этом гипотеза Бернштейна оказалась верной. После смены знака мембранного потенциала ионы натрия продолжают перемещаться в клетку только под действием концентрационного градиента. Мембранный потенциал на максимуме овершута приближается к равновесному электрохимическому потенциалу для ионов
внутри клетки примерно в десять раз ниже чем в межклеточной жидкости. В начале спайка ионы натрия под действием электрического поля и концентрационного градиента устремляются из межклеточной среды в клетку (в клетке их концентрация ниже, а внутренняя поверхность мембраны заряжена отрицательно по отношению к внешней). В этом гипотеза Бернштейна оказалась верной. После смены знака мембранного потенциала ионы натрия продолжают перемещаться в клетку только под действием концентрационного градиента. Мембранный потенциал на максимуме овершута приближается к равновесному электрохимическому потенциалу для ионов ![]() . В дальнейшем проницаемость мембраны для ионов натрия падает и активируется проводимость для ионов калия, которые движутся из клетки, где их концентрация выше. Ионы калия уносят из клетки положительный заряд. Внутренняя поверхность мембраны заряжается отрицательно. Этот натриево-калиевый цикл ниже будет рассмотрен подробнее.
. В дальнейшем проницаемость мембраны для ионов натрия падает и активируется проводимость для ионов калия, которые движутся из клетки, где их концентрация выше. Ионы калия уносят из клетки положительный заряд. Внутренняя поверхность мембраны заряжается отрицательно. Этот натриево-калиевый цикл ниже будет рассмотрен подробнее.
В течение цикла ионы натрия заходят в клетку, а ионы калия ее покидают. Такое замещение должно выравнивать концентрации ионов. Казалось бы, что после ряда циклов нервная клетка не должна генерировать спайков (концентрации ионов внутри и вне клетки уравняются). В действительности это не так. Неравномерное распределение ионов внутри и вне клетки поддерживается активным транспортом ионов. Механизмы, осуществляющие транспорт ионов через мембрану в сторону повышенной концентрации, называются мембранными насосами (помпами). Они потребляют энергию, которая освобождается в результате окисления аминокислот.
Наиболее изучен натриевый насос. Он выводит ионы ![]() из клетки и одновременно транспортирует ионы
из клетки и одновременно транспортирует ионы ![]() внутрь клетки. Этот процесс включает в себя обязательный обмен двух ионов
внутрь клетки. Этот процесс включает в себя обязательный обмен двух ионов ![]() во внешней среде на три иона
во внешней среде на три иона ![]() в клетке (если удалить из среды ионы
в клетке (если удалить из среды ионы ![]() , то ионы
, то ионы ![]() из клетки выводиться уже не будут). Если мембранный потенциал отрицателен (внутренняя поверхность мембраны заряжена отрицательно), то натриевый насос выполняет электрическую работу, выводя из клетки суммарно один положительный заряд. Натриевый насос действует избирательно. Например, он не способен переносить ионы лития. Молекулярный механизм активного транспорта в настоящее время до конца не ясен.
из клетки выводиться уже не будут). Если мембранный потенциал отрицателен (внутренняя поверхность мембраны заряжена отрицательно), то натриевый насос выполняет электрическую работу, выводя из клетки суммарно один положительный заряд. Натриевый насос действует избирательно. Например, он не способен переносить ионы лития. Молекулярный механизм активного транспорта в настоящее время до конца не ясен.