Математические модели естествознанияРефераты >> Естествознание >> Математические модели естествознания
Электрохимическое уравнение (3) суть обыкновенное дифференциальное уравнение первого порядка. Как показано выше, его коэффициенты следует считать функциями от мембранного потенциала ![]() . По биологическому смыслу решения уравнения ограничены на положительной полуоси
. По биологическому смыслу решения уравнения ограничены на положительной полуоси ![]() . Можно доказать, что все решения такого уравнения монотонно стремятся к состояниям равновесия. Объяснить в рамках одного электрохимического уравнения генерацию потенциала действия невозможно.
. Можно доказать, что все решения такого уравнения монотонно стремятся к состояниям равновесия. Объяснить в рамках одного электрохимического уравнения генерацию потенциала действия невозможно.
Из приведенных выше построений следует, проводимости ![]() и
и ![]() следует считать функциями не только мембранного потенциала
следует считать функциями не только мембранного потенциала ![]() , но и времени. Нижеследующие построения А. Ходжкина и А. Хаксли во многом носят феноменологический характер, т.е. не имеют под собой прочной теоретической основы и опираются на ряд допущений и догадок. Рассмотрим натриевую проводимость. Для описания ее зависимости от времени и мембранного потенциала А. Ходжкин и А. Хаксли ввели две новые переменные
, но и времени. Нижеследующие построения А. Ходжкина и А. Хаксли во многом носят феноменологический характер, т.е. не имеют под собой прочной теоретической основы и опираются на ряд допущений и догадок. Рассмотрим натриевую проводимость. Для описания ее зависимости от времени и мембранного потенциала А. Ходжкин и А. Хаксли ввели две новые переменные ![]() и
и ![]() , которые характеризуют процессы активации и инактивации натриевых каналов. В интерпретации авторов
, которые характеризуют процессы активации и инактивации натриевых каналов. В интерпретации авторов ![]() -вероятность открытия
-вероятность открытия ![]() -ворот,
-ворот, ![]() -вероятность закрытия
-вероятность закрытия ![]() -ворот, или же -отношение числа открытых и закрытых ворот соответствующего типа к числу каналов на поверхности мембраны единичной площади. Авторы модели положили:
-ворот, или же -отношение числа открытых и закрытых ворот соответствующего типа к числу каналов на поверхности мембраны единичной площади. Авторы модели положили: ![]() , где коэффициент
, где коэффициент ![]() был назван максимальной натриевой проводимостью. Предложенное А. Ходжкиным и А. Хаксли обоснование вида зависимости в настоящее время выглядит искусственным. Методом подбора были выписаны линейные обыкновенные уравнения первого порядка для переменных
был назван максимальной натриевой проводимостью. Предложенное А. Ходжкиным и А. Хаксли обоснование вида зависимости в настоящее время выглядит искусственным. Методом подбора были выписаны линейные обыкновенные уравнения первого порядка для переменных ![]() и
и ![]() . При этом в уравнение для
. При этом в уравнение для ![]() не входит переменная
не входит переменная ![]() , а в уравнение для
, а в уравнение для ![]() не входит
не входит ![]() . Коэффициенты уравнений зависят только от мембранного потенциала
. Коэффициенты уравнений зависят только от мембранного потенциала ![]() . Более менее убедительное обоснование необходимости именно такой зависимости отсутствует.
. Более менее убедительное обоснование необходимости именно такой зависимости отсутствует.
Для описания изменения калиевой проводимости ![]() А. Ходжкиным и А. Хаксли была введена функция
А. Ходжкиным и А. Хаксли была введена функция ![]() . Она интерпретировалась как вероятность открытия
. Она интерпретировалась как вероятность открытия ![]() -ворот для калиевых каналов, т.е. как вероятность активации последних. Напомним, что по гипотезе А. Ходжкина и А. Хаксли инактивационные ворота для калиевых каналов отсутствуют. Авторы опытным путем подобрали линейное обыкновенное дифференциальное уравнение для
-ворот для калиевых каналов, т.е. как вероятность активации последних. Напомним, что по гипотезе А. Ходжкина и А. Хаксли инактивационные ворота для калиевых каналов отсутствуют. Авторы опытным путем подобрали линейное обыкновенное дифференциальное уравнение для ![]() и положили
и положили ![]() . Коэффициенты уравнения зависят только от мембранного потенциала
. Коэффициенты уравнения зависят только от мембранного потенциала ![]() . Коэффициент
. Коэффициент ![]() называется максимальной калиевой проводимостью.
называется максимальной калиевой проводимостью.
Относительно проводимости ![]() для тока утечки А. Ходжкин и А. Хаксли предложили считать его постоянным:
для тока утечки А. Ходжкин и А. Хаксли предложили считать его постоянным: ![]() . В результате для описания процесса генерации потенциала действия была предложена система четырех уравнений, Первое из них -электрохимическое уравнение для баланса мембранных токов :
. В результате для описания процесса генерации потенциала действия была предложена система четырех уравнений, Первое из них -электрохимическое уравнение для баланса мембранных токов : ![]() . Напомним, что
. Напомним, что
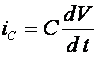 ,
, ![]() , (4)
, (4)
![]() ,
, ![]() . (5)
. (5)
Второе и третье уравнения описывают соответственно процессы открытия ![]() и закрытия
и закрытия ![]() -ворот для натриевых каналов. Наконец, четвертое уравнение отражает процесс открытия
-ворот для натриевых каналов. Наконец, четвертое уравнение отражает процесс открытия ![]() -ворот для калиевых каналов. Система уравнений Ходжкина -Хаксли имеет следующий вид:
-ворот для калиевых каналов. Система уравнений Ходжкина -Хаксли имеет следующий вид:
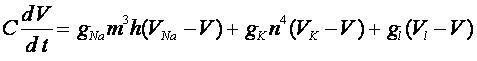 , (6)
, (6)
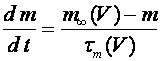 , (7)
, (7)
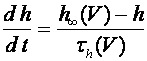 , (8)
, (8)
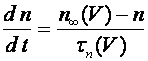 , (9)
, (9)
где, напомним, ![]() ,
, ![]() ,
, ![]() . В (7) -(9) функции
. В (7) -(9) функции ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() положительны и непрерывны. Функции
положительны и непрерывны. Функции ![]() ,
, ![]() ,
, ![]() относятся к классу так называемых сигмоидных функций, т.е. они монотонны обладают следующими свойствами:
относятся к классу так называемых сигмоидных функций, т.е. они монотонны обладают следующими свойствами: ![]() ,
, ![]() ,
, ![]() при
при ![]() и
и ![]() ,
, ![]() ,
, ![]() при
при ![]() . В работах А. Ходжкина и А. Хаксли указан их конкретный вид. Функции весьма сложно устроены. Реально система уравнений допускает только численное исследование. Ограничимся рядом замечаний на качественном уровне, иллюстрирующих впечатления от численного исследования. Входящие в систему (6) -(9) функции подобраны так, что выполнен ряд свойств. Она имеет устойчивое состояние равновесия
. В работах А. Ходжкина и А. Хаксли указан их конкретный вид. Функции весьма сложно устроены. Реально система уравнений допускает только численное исследование. Ограничимся рядом замечаний на качественном уровне, иллюстрирующих впечатления от численного исследования. Входящие в систему (6) -(9) функции подобраны так, что выполнен ряд свойств. Она имеет устойчивое состояние равновесия ![]() , где
, где ![]() ,
, ![]() -положительны. Пусть
-положительны. Пусть ![]() ,
, ![]() ,
, ![]() ,
, ![]() . Если значение
. Если значение ![]() не слишком велико, то в силу устойчивости состояния равновесия с течением времени
не слишком велико, то в силу устойчивости состояния равновесия с течением времени ![]() . Существует такое пороговое значение
. Существует такое пороговое значение ![]() , что при
, что при ![]() решения системы ведут себя по другому. Первоначально
решения системы ведут себя по другому. Первоначально ![]() убывает. Однако начинает расти величина
убывает. Однако начинает расти величина ![]() . Слагаемое
. Слагаемое ![]() уравнении (6) положительно, т.к.
уравнении (6) положительно, т.к. ![]() , а
, а ![]() близко к нулю. В результате производная V¢(t) становится положительной. Мембранный потенциал начинает расти (момент начала генерации спайка). Вместе с ним растет и величина
близко к нулю. В результате производная V¢(t) становится положительной. Мембранный потенциал начинает расти (момент начала генерации спайка). Вместе с ним растет и величина ![]() , что приводит к дальнейшему увеличению V¢(t) и ускорению роста
, что приводит к дальнейшему увеличению V¢(t) и ускорению роста ![]() . Однако, чуть позже начинает уменьшаться величина
. Однако, чуть позже начинает уменьшаться величина ![]() . Это замедляет рост
. Это замедляет рост ![]() , тем более, что уменьшается разность
, тем более, что уменьшается разность ![]() . Одновременно растет величина
. Одновременно растет величина ![]() . Соответствующее слагаемое
. Соответствующее слагаемое ![]() в уравнении (6) отрицательно, т.к.
в уравнении (6) отрицательно, т.к. ![]() и
и ![]() . Существует момент времени
. Существует момент времени ![]() , где V¢(tm)=0. Это точка максимума потенциала действия. Значение мембранного потенциала
, где V¢(tm)=0. Это точка максимума потенциала действия. Значение мембранного потенциала ![]() относительно близко к
относительно близко к ![]() . После прохождения точки максимума значение производной V¢(t) определяется значением слагаемого
. После прохождения точки максимума значение производной V¢(t) определяется значением слагаемого ![]() и становится отрицательным. Скорость убывания мембранного потенциала на нисходящем участке спайка меньше по абсолютной величине, чем скорость роста на восходящем участке. Нисходящий участок продолжительнее восходящего. В дальнейшем значение
и становится отрицательным. Скорость убывания мембранного потенциала на нисходящем участке спайка меньше по абсолютной величине, чем скорость роста на восходящем участке. Нисходящий участок продолжительнее восходящего. В дальнейшем значение ![]() становится отрицательным, а после этого
становится отрицательным, а после этого ![]() . Величины
. Величины ![]() ,
, ![]() ,
, ![]() также стремятся к своим равновесным значениям. Процесс генерации спайка закончен.
также стремятся к своим равновесным значениям. Процесс генерации спайка закончен.