Апоптоз - программируемая клеточная смерть
Апоптоз – многоэтапный процесс. Первый этап – прием сигнала, предвестника гибели в виде информации, поступающей к клетке извне или возникающей в недрах самой клетки. Сигнал воспринимается рецептором и подвергается анализу.
Далее через рецепторы или их сочетания полученный сигнал последовательно передается молекулам-посредникам (мессенджерам) различного порядка и в конечном итоге достигает ядра, где и происходит включение программы клеточного самоубийства путем активации летальных и/или репрессии антилетальных генов. Однако существование ПКС (программируемая клеточная смерть) в безъядерных системах (цитопластах – клетках, лишенных ядра) показывает, что наличие ядра не является обязательным для реализации процесса ].
Применительно к клеткам животных и человека апоптоз в большинстве случаев связан с протеолитической активацией каскада каспаз – семейства эволюционно консервативных цистеиновых протеаз, которые специфически расщепляют белки после остатков аспарагиновой кислоты .
На основе структурной гомологии каспазы подразделяются на подсемейства
а) каспазы-1 (каспазы 1, 4, 5),
б) каспазы-2 (каспаза-2) и
в) каспазы-3 (каспазы 3, 6–10) .
Цистеиновые протеазы, по-видимому, участвуют также в ПКС у растений . Однако апоптоз возможен и без участия каспаз: сверхсинтез белков-промоторов апоптоза Bax и Bak индуцирует ПКС в присутствии ингибиторов каспаз .
В результате действия каспаз происходит:
1.активация прокаспаз с образованием каспаз;
2.расщепление антиапоптозных белков семейства Bcl-2. Подвергается протеолизу ингибитор ДНКазы, ответственный за фрагментацию ДНК. В нормальных клетках апоптозная ДНКаза CAD (caspase-activated DNase) образует неактивный комплекс с ингибитором CAD, обозначаемым ICAD или . При апоптозе ингибитор ICAD с участием каспаз 3 или 7 инактивируется , и свободная CAD, вызывая межнуклеосомальные разрывы хроматина, ведет к образованию фрагментов ДНК с молекулярной массой, кратной молекулярной массе ДНК в нуклеосомных частицах – 180-200 пар нуклеотидов.
Апоптоз возможен и без фрагментации ДНК . Обнаружен ядерный белок Acinus (apoptotic chromatin condensation inducer in the nucleus), из которого при комбинированном действии каспазы-3 (протеолиз при Asp 1093) и неидентифицированной протеазы (протеолиз при Ser 987) образуется фрагмент Ser 987 – Asp 1093. Этот фрагмент в присутствии дополнительных неядерных факторов вызывает апоптотическую конденсацию хроматина и фрагментацию ядра (кариорексис) без фрагментации ДНК ;
3.гидролиз белков ламинов, армирующих ядерную мембрану. Это ведет к конденсации хроматина;
4.разрушение белков, участвующих в регуляции цитоскелета;
5.инактивация и нарушение регуляции белков, участвующих в репарации ДНК, сплайсинге мРНК, репликации ДНК.
(((Мишенью каспаз является поли(ADP-рибозо)полимераза (ПАРП). Этот фермент участвует в репарации ДНК, катализируя поли(ADP-рибозилирование) белков, связанных с ДНК (см. обзоры [3,11]). Донором ADP-рибозы является NAD+. Активность ПАРП возрастает в 500 раз и более при связывании с участками разрыва ДНК. Апоптотическая гибель клетки сопровождается расщеплением ПАРП каспазами. Чрезмерная активация ПАРП при массированных разрывах ДНК, сильно снижая содержание внутриклеточного NAD+, ведет к подавлению гликолиза и митохондриального дыхания и вызывает гибель клетки по варианту некроза. ))))
Существует несколько путей реализации программы ПКС .
Среди них важное место занимает путь, опосредованный физиологическими индукторами, действие которых реализуется через клеточные рецепторы, специально предназначенные для включения программы апоптоза. Этот путь передачи сигнала ПКС схематически можно изобразить следующим образом: индукторы ’ рецепторы ’ адаптеры ’ каспазы первого эшелона ’ регуляторы ’ каспазы второго эшелона. Так, рецептор, обозначаемый Fas, взаимодействуя с соответствующим лигандом (лигандом FasL), трансмембранным белком Т-киллера, активируется и запускает программу смерти клетки, инфицированной вирусом. Тем же путем при взаимодействии с лигандом FasL на поверхности ТН-1-лимфоцитов или с антителом к Fas-рецептору погибают ставшие ненужными выздоровевшему организму В-лимфоциты, продуценты антител, несущие Fas-рецептор. FasL– лиганд, относящийся к многочисленному семейству фактора некроза опухолей TNF. Это семейство гомотримерных лигандов, кроме FasL и TNFa , включает TNFb (лимфотоксин).
Fas – член семейства рецепторов TNF. Все они представлены трансмембранными белками, которые внеклеточными участками взаимодействуют с тримерами лигандов-индукторов . Взаимодействие рецептора и лиганда приводит к образованию кластеров рецепторных молекул и связыванию их внутриклеточных участков с адаптерами. Адаптер, связавшись с рецептором, вступает во взаимодействие с эффекторами, пока еще неактивными предшественниками протеаз из семейства каспаз первого эшелона (инициирующих каспаз).
Взаимодействие адаптера с рецептором и эффектором осуществляется через гомофильные белок-белковые взаимодействия небольших доменов: DD (death domain – домен смерти), DED (death-effector domain – домен эффектора смерти), CARD (– домен активации и рекрутирования каспазы). Все они имеют сходную структуру, содержат по шесть a-спиральных участков. Домены DD(домен смерти) участвуют во взаимодействии рецептора Fas c адаптером FADD (Fas-associated DD-protein). Домены DED участвуют во взаимодействии адаптера FADD с прокаспазами 8 и 10.
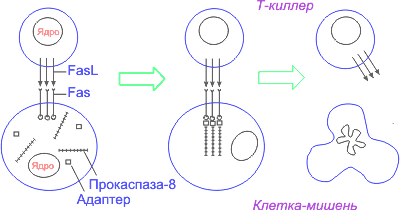
Рис. 2. Зависимый от Fas-рецептора апоптоз клетки-мишени при действии цитотоксического Т-лимфоцита (Т-киллера)
Наиболее подробно охарактеризована прокаспаза-8, рекрутируемая рецептором Fas через адаптeр FADD. Образуются агрегаты FasL – Fas – FADD – прокаспаза-8. Подобные агрегаты, в которых происходит активация каспаз, названы апоптосомами , апоптозными шаперонами , или сигнальными комплексами, индуцирующими смерть.
Прокаспазы обладают незначительной протеолитической активностью, составляющей 1–2% активности зрелой каспазы. Будучи в мономерной форме, прокаспазы, концентрация которых в клетке ничтожна, находятся в латентном состоянии. Предполагается, что пространственное сближение молекул прокaспаз при их агрегации ведет к образованию активных каспаз через механизм протеолитического само- и перекрестного расщепления (ауто- или транс-процессинга)]. В результате от прокаспазы (молекулярная масса 30–50 кДа) отделяется регуляторный N-концевой домен (продомен), а оставшаяся часть молекулы разделяется на большую (~20 кДа) и малую (~10 кДа) субъединицы (рис. 3). Затем происходит ассоциация большой и малой субъединиц. Два гетеродимера образуют тетрамер с двумя каталитическими участками, действующими независимо друг от друга. Таким образом прокаспаза-8 активируется и высвобождается в цитоплазму в виде каспазы-8. Существуют другие пути активации каспазы-8 – с участием рецепторов TNFR1 и DR3.
На этапе активации каспаз первого эшелона жизнь клетки еще можно сохранить. Существуют регуляторы, которые блокируют или, напротив, усиливают разрушительное действие каспаз первого эшелона. К ним относятся белки Bcl-2 (ингибиторы апоптоза: A1, Bcl-2, Bcl-W, Bcl-XL, Brag-1, Mcl-1 и NR13) и Bax (промоторы апоптоза: Bad, Bak, Bax, Bcl-XS, Bid, Bik, Bim, Hrk, Mtd). Эти белки эволюционно консервативны: гомолог Bcl-2 обнаружен даже у губок, у которых апоптоз необходим для морфогенеза .