Взаимосвязь основных внутриколониальных процессов при ветвлении у колониальных гидроидов
Колониальные гидроиды стали удачной моделью для изучения механизмов интеграции примитивных биосистем организменного уровня. В процессе роста форма, размеры и число компонентов колонии все время меняются, но остаются оптимальными для данных условий существования. Это достигается с помощью регуляции потребления питательного материала при линейном росте, ветвлении и увеличении объема тела, связанных друг с другом определенной последовательностью смены процессов.
Особенности морфологии и физиологии гидроидных полипов делают их одними из наиболее удачных объектов для исследования закономерностей взаимосвязи отдельных морфогенетических актов с единым процессом формообразования целостной, хорошо интегрированной живой системы — колонии гидроидов.
Все части колонии морфологически и по происхождению являются единым телом с общей гастроваскулярной полостью, через которую осуществляется транспорт пищи, продуктов обмена веществ и, возможно, клеток на значительные расстояния. Таким образом, взаимосвязь основных внутриколониальных процессов (пищеварения, пролиферации, роста, редукции частей колонии, ветвления) осуществляется через гидроплазму—жидкость, заполняющую полость колонии. Несмотря на примитивность распределительной системы, ее функционирование обеспечивает достаточно эффективную физиологическую интеграцию колонии в единый организм (Карлсен, Марфенин, 1984; Марфенин, 1985, 1986). Это проявляется в поддержании в процессе роста колонии определенных соотношений между количествами разнотипных составляющих. Нарушение количественных пропорций вызывает такие изменения в росте, которые приводят к восстановлению утраченных соотношений (Марфенин, 1977; Stebbing. 198I).
В настоящее время уже получены данные о характере зависимости роста колонии от интенсивности питания (Bravennan, 197I, 1974; Марфенин, Бурыкин, 1979; Бурыкин, 1980; Марфенин, 1987), о степени автономности ростовых пульсаций верхушек столонов и побегов (Косевич, 1988).
Назрела необходимость ответить на вопрос: от каких факторов зависит ветвление колоний, играющее ведущую роль в обеспечении их оптимального строения и роста? Решая эту задачу, мы использовали оправдавший себя ранее метод анализа морфологии объекта при разных режимах его кормления (Марфенин, 1986) и удобный модельный объект — стелющиеся колонии гидроида Cordylophora inkermanica (Marfenin, 1983).
Материал и методы
С. inkermanica — представитель примитивных колоний стелющегося типа из сем. Clavidae, подотряда Athecata, отряда Leptolida. У этих гидроидов побеги короткие и ветвятся слабо, поэтому зооиды отходят обычно непосредственно от столонов. Материал собран в Севастопольской бухте Черного моря; гидроидов культивировали в лаборатории. Морфология и биология вида описаны ранее (Марфенин, 1983, 1985). Проведены три серии экспериментов: первая и вторая—на Беломорской биостанции МГУ в 1984 и 1987 гг., третья—на кафедре зоологии беспозвоночных биологического факультета МГУ в 1984 г. В первых двух сериях животных содержали в 5-литровом кристаллизаторе, воду в котором меняли один раз в два дня. В третьей серии опытов гидроиды находились в 40-литровом аквариуме с беломорской водой (соленость 28°/оо). Осуществлялась постоянная аэрация и фильтрация воды через отдельный грунтовый фильтр. Воду не меняли 4 мес. Температура содержания в первой серии была 14±2°С, во второй 15±2°С и в третьей 14±1°С.
Колонии выращивали на стеклах по методике Кроуэлла (Crowell,, 1957) из отрезков столона длиной 1,5—4,5 мм, в каждом из которых было по одному гидранту. Гидрант и отрезок мы называем “начальными”, а столон, являющийся продолжением начального отрезка,— “центральным” или “главным” столоном. На центральном столоне в результате его ветвления образуются боковые столоны (рис. 1, 2).
Эксперимент проводили в первые 8—14 сут роста начального отрезка. Каждый гидрант колонии кормили индивидуально из пипетки. Кормом служили свежевылупившиеся науплиусы Artemia sp. Среднесуточное количество корма, полученное каждой колонией, было пропорционально ее размеру, который выражали суммарной длиной гидроризы в миллиметрах. Удельные количества пищи значительно различались в колониях, что позволяло сравнивать эффект кормления. Ежедневно проводили картирование колоний по известной методике (Марфенин, 1980). Всего был изучен рост 66 колоний. Для определения объемных параметров роста в 14 из них проведено измерение диаметров перисарка, ценосарка и гастральной полости. Измерения проводили in vivo в проходящем свете под микроскопом с точностью до 4 мкм через каждые 0,5 мм по длине столона. Таким образом, число промеров было пропорционально размерам колоний. В малых колониях (с длиной гидроризы 8—9 мм) сделано по 14—18, а в больших (длиной 80—100 мм) по 150—200 промеров. По результатам измерений вычислены значения толщины стенок ценосарка столонов, что дало возможность определить объем ткани в них.
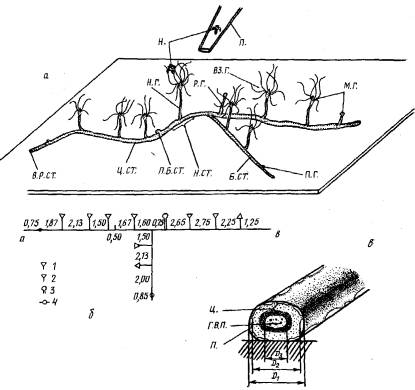
Рис.1. a — общий вид небольшой колонии, выращенной на пластине из оргстекла. Н. СТ.—начальный столон; Н. Г.—начальный гидрант; Ц, СТ.—центральный столона Б. СТ.— боковой столон; П. Б. СТ.— почка бокового столона; В. Р. ОТ.— верхушка роста столона; П. Г.— почка гидранта; М. Г.— молодой гидрант; ВЗ. Г.— взрослый гидрант; Р. Г.— редуцирующийся гидрант; Н — науплиус Artemia sp.; П.— кончик пипетки; б—карта-схема этой же колонии. Цифры—расстояние между структурами, мм:
1—взрослый гидрант; 3—молодой гидрант; 3—редуцирующийся гидрант; 4—почка гидранта; в — схема поперечного среза столона. Промеры диаметров: D1 — перисарка (П.); D2.—ценосарка (II.); D3—гастроваскулярной полости (Г. В. П.)
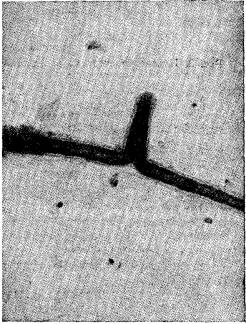
Рис. 2. Фотография бокового столона через 12 ч после его образования
РЕЗУЛЬТАТЫ
Поскольку колонии существенно различны по размерам, то абсолютное число науплиусов (F1), полученное каждой из них в течение эксперимента, не может служить показателем интенсивности питания.
Универсальной характеристикой размера колонии является длина гидроризы (L). Остальные параметры — число верхушек роста столонов (В), число гидрантов—редуцирующихся (Hd), формирующихся (Нi2) и взрослых, сформированных (H),—пропорционально L.
Поэтому интенсивность питания мы определяли удельным среднесуточным количеством пищи F/Lt; (науплиусов/(мм-сут), где t—период времени, сут, который прошел со времени выделения начального отрезка до того, как проводили измерения. Интенсивность питания в разных колониях была различна—от 0,03 до 0,49 науплиусов/(мм-сут).
Динамика роста существенно различалась в колониях, получавших различное количество пищи. Для характеристики морфологии и динамики роста колоний мы использовали следующие показатели.
1. Скорость роста верхушек столона (l/В) отражает величину средней скорости роста любого столона в колонии. Для его вычисления значение суточного прироста всей колонии делили на общее число верхушек ее столонов.