Взаимосвязь основных внутриколониальных процессов при ветвлении у колониальных гидроидов
С увеличением удельного среднесуточного количества пищи возрастает средняя площадь сечения трубки ценосарка столона (рис. 8). Зависимость площади сечения перисарка от питания имеет аналогичный характер. В среднем площадь сечения ценосарка в экспериментальных колониях составил 2,83×10-3 мм3. Минимальное значение, которое было зарегистрировано,—1,82×10-3, а максимальное — 4,43×10-3 мм3. Разность объемов ткани ценосарка двух колоний одинаковой длины (L=20 мм), в одной из которых интенсивность питания была 0,050 науплиусов/(мм×сут), а в другой 0,380 науплиусов/(мм×сут), составила 1,2×10-2 мм3. Эта величина соответствует объему ценосарка столона длиной 4 мм, т. е. трехдневному приросту одной верхушки столона при обильном питании (0,300 науплиусов/(мм×сут). Расчеты показали, что если удельное среднесуточное количество пищи в одной колонии на 0,400 науплиусов/(мм×сут) больше), чем в другой, то общие объемы тканей ценосарков этих колоний должны различаться в 2 раза. Кроме того, подсчитано, что возрастание интенсивности питания на 0,050 пауплиусов/(мм×сут) вызывает увеличение объема ткани, эквивалентное объему ценосарка столона длиной 0,6 мм. Скорость роста столона при таком изменении интенсивности питания увеличивается на 1 мм/сут.
При недостаточном питании (менее 0,100 науплиусов/(мм×сут)) в колонии происходит редукция столонов и гидрантов. Ценосарк при этом уменьшается в толщине — “худеет” и столон укорачивается с концов. Число редуцирующихся гидрантов в таких колониях превосходит число растущих. На приведенных графиках этой ситуации соответствуют отрицательные значения показателей 1/В, В—2/L и HgfL (рис. 4, 6, 7).
). 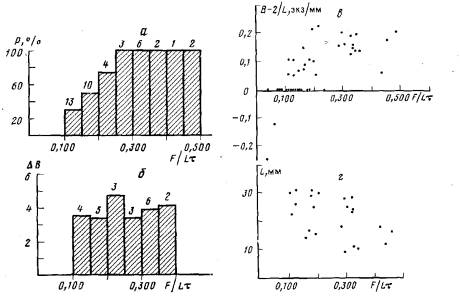
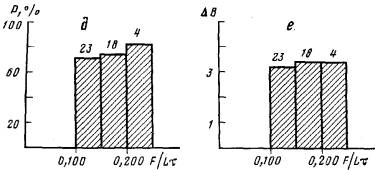
Рис. 6. Влияние литания на ветвление гидроризы колонии. Цифры над гистограммами указывают объем выборок, а—зависимость вероятности образования почек боковых столонов (Р) от интенсивности питания (F/Lr) при равномерном кормлении; б—зависимость числа почек боковых столонов (В) от интенсивности питания (F/Lt) при равномерном кормлении; в — изменение показателя раэветвленности гидроризы (В — 2/L) в зависимости от интенсивности питания {F/Lt} при равномерном кормлении; г — значения интенсивности питания н размера колонии (L), при котором произошло ветвление в колониях при равномерном кормлении; д—зависимость вероятности образования почек боковых столонов (Р) от интенсивности питания {F/Lt} при импульсном кормлении; е—зависимость числа почек боковых столонов (6) от интенсивности питания {F/Lt} при импульсном кормлении
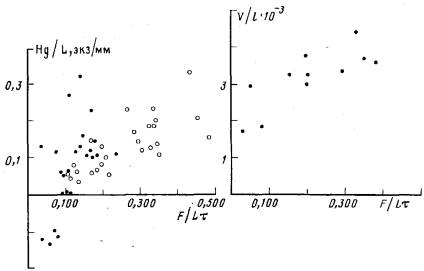
Рис. 7. Зависимость удельного числа гидрантов, на рост которых расходуется пища (Hg/L), от интенсивности питания(F/Lt)
Рис, 8. Зависимость удельного объема ткани ценосарка столона (V/L) от интенсивности питания {F/Lt)
Обсуждение результатов
Изучение закономерностей ветвления в колонии гидроидных полипов имеет большое значение для выяснения регуляторных механизмов поддержания такой структуры колонии, которая наилучшим образом соответствовала бы условиям ее существования, и прежде всего трофической емкости среды. Ранее было показано (Марфенин, 1977), что в колонии гидроидов в процессе ее роста сохраняется определенная пропорция между числом гидрантов, числом верхушек роста столонов и длиной гидроризы. Это способствует наиболее эффективному усвоению пищи, в результате которого обеспечивается быстрый рост колонии. Было установлено, что соответствие между ростом и его трофическим обеспечением осуществляется посредством четырех основных механизмов.
1. Изменение скорости роста столонов в определенном диапазоне в зависимости от интенсивности питания было зарегистрировано Ю. Б. Бурыкиным (1980) на колониальном гидроиде Dynamena pumila L. В дальнейшем эта зависимость была подтверждена на другом Obelia loveni Allm. (Марфенин, 1987). При увеличении скорости роста столонов происходит не только удлинение их тела, но и увеличение числа новообразованных гидрантов или побегов, каждый из которых также растет, а следовательно, потребляет клеточный материал. Поэтому возможность изменения в некоторых пределах скорости роста столонов позволяет нелинейно менять интенсивность потребления в колонии клеточного материала.
2. Изменение доли растущих побегов и столонов было впервые обнаружено Н. Н. Марфениным (1977) также при изучении D. pumila. Практически всегда в колонии растут не все имеющиеся верхушки роста. Доля растущих от общего числа имеющихся в наличии может существенно меняться — при недостатке пищи часть верхушек побегов и столонов прекращает расти, а при избытке пищи часть нерастущих верхушек возобновляет свой рост.
3. Ткани редуцирующихся гидрантов служат дополнительным источником пищи. Лангер (Lunger, 1963) называет ее “вторичной пищей”. С помощью оптической, а затем и электронной микроскопии было показано, что клетки дедифференцирующихся гидрантов попадают в гастроваскулярную полость побегов (Huxley, De Beer, 1923) и гидроризы (Полтева и др., 1987). Затем энтодермальные клетки гидроризы фагоцитируют и переваривают частицы вторичной пищи (Lunger, 1963). Таким образом, существуют как бы два типа источников пищи для колонии. Первый — внешняя среда, пищевые ресурсы которой потенциально не ограничены; второй — дедифференцирующиеся части самой колонии. Количество питательного материала, поступающего из этого источника, ограничено объемом тканей редуцирующихся частей колонии.
4. Образование новых зон роста или ветвление колонии происходит как в процессе обычного роста столонов (новые гидранты), так и при избытке пищи (Марфенин, Бурыкин, 1979; Бурыкин, 1980). Ветвление не сдерживается какими-либо рамками и может привести к почти неограниченному увеличению числа новых зон роста. Поэтому этот способ регуляции роста колонии был признан наиболее радикальным (Марфенин, 1977).
Если образование очередных побегов или гидрантов (в случае стелющейся колонии) происходит регулярно по мере роста столонов примерно на равных расстояниях друг от друга, то образование боковых столонов выглядит неупорядоченно: ни место, ни число новых столонов не бывают точно определены. Однако всякий раз, когда происходит ветвление, оно приводит к нормализации количественного баланса между различными компонентами колонии—нормализации, имеющей большое экологическое значение” так как в результате достигается соответствие между ростом колонии и трофическими возможностями среды.
Полученные данные позволяют представить, как работает этот механизм. Образование новых зон роста—ветвление столонов—связано не только с интенсивностью питания колонии, но и со скоростями роста имеющихся верхушек, толщиной трубки ценосарка, количеством растущих гидрантов и интенсивностью редуцирования старых гидрантов.
Все эти внутриколониальные процессы взаимосвязаны через потребление питательного материала.