Влияние повышенного и сниженного уровня моноаминов на функциональную организацию колонок C1 коры мозга крысы
В выборке попарно зарегистрированных нейронов (N=28) оценивались влияния активации ядер шва на характер межнейронных взаимодействий с использованием метода построения ПИГ. Значимые пики на ПИГ рассматривали как проявление статистически выявленной функциональной связи между нейронами, осуществляющейся с определенной задержкой и в определенном направлении.
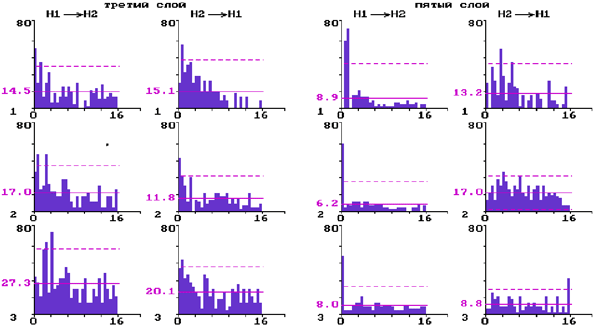
Рис.2. Перестройки межнейронных взаимодействий нейронов на разных уровнях бочонковой колонки С1 коры мозга крыс при электрической стимуляции ядер шва.
Постимпульсные гистограммы связи для прямых и обратных влияний парных нейронов слоя 3 и парных нейронов слоя 5:
1 - до стимуляции ядер шва; 2 - через десять минут; 3 - через двадцать минут после стимуляции. По вертикали - количество импульсов; по горизонтали - время в мс. Сплошная горизонтальная линия - среднее количество импульсов; штриховая - уровень 2s .
Для верхних слоев показано (рис. 2) появление на ПИГ после электрической стимуляции дополнительных длиннолатентных значимых пиков и уменьшение пиков с нулевой латентностью. В нижних слоях при этом наблюдалось ослабление как прямых, так и обратных межнейронных моно- и полисинаптических влияний и появление односторонних влияний с нулевой и высокой латентностью.
Сниженное под влиянием резерпина внутримозговое содержание СТ.
Анализ средней текущей частоты фоновой импульсной активности нейронов бочонковой колонки С1 до и после введения резерпина, результаты которого представлены в таблице, показал, что на пике действия резерпина суммарно по колонке, и на каждом из ее уровней имеет место достоверное (Р<0,05) снижение частоты импульсации. При этом характерный для контроля более высокий частотный уровень нейронов поверхностных слоев по сравнению с глубокими слоями на фоне резерпина выравнивался. Следует отметить, что около 18 % нейронов, исходная частота импульсации которых не превышала средних значений по выборке (Р<0,05), были резистентными к резерпину. У 10 % нейронов, которые зарегистрированы преимущественно в эфферентных слоях, частота импульсации в последействии резерпина достоверно (Р<0,05) повышалась. Их исходная частота превышала среднее значение по всей выборке (Р<0,01).
На рис. 3 представлены ГМИ фоновой импульсной активности трех нейронов, принадлежащих бочонковой колонке С3. Нейрон на (А) имел до введения резерпина три отчетливых максимума в ГМИ, что свидетельствует о наличии в структуре его исходного разряда пачек и групп, перемежающих аритмично следующие единичные разряды. Через 2,5 ч после введения резерпина частота импульсации
данного нейрона уменьшилась, а структура рандомизировалась - исчезли пачечные и групповые разряды. Средний межимпульсный интервал у снижающих частоту нейронов (N=10) в контроле был равен 19,8± 4,4 мс, а через 2-3 часа после введения резерпина - 24,5± 2,6 мс, что по t- критерию недостоверно отличается от контроля (P<0,05).
У нейрона на (Б) через 2 ч после введения резерпина ГМИ стала островершинной с двумя достоверными максимумами в диапазоне самых малых интервалов, что отражает появление в структуре непрерывного аритмичного разряда коротких пачек и разной длительности групп импульсов. Такая динамика структуры фонового разряда была характерна для нейронов, которые увеличивали под влиянием резерпина частоту импульсной активности. Средний межимпульсный интервал у повышающих частоту нейронов (N=10) в контроле был равен 22,4± 6,6 мс, а через 2-3 часа после введения резерпина - 19,6± 3,9 мс, что по t- критерию недостоверно отличается от контроля (P<0,05).
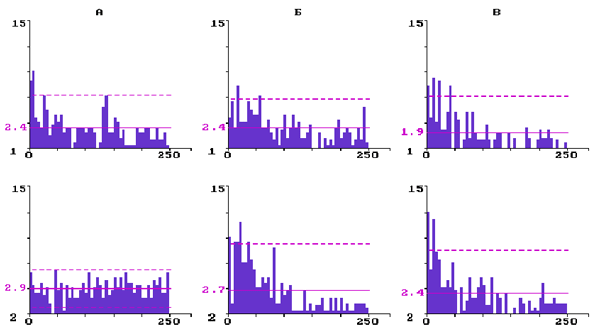
Рис.3. Перестройка структуры фоновой импульсной активности трех нейронов разных уровней бочонковой колонки С1 коры мозга крыс при действии резерпина
Гистограммы межимпульсных интервалов: А - при уменьшении частоты импульсации; Б - при увеличении частоты; В - при стабильной частоте; 1 - в контроле; 2 - в последействии резерпина. По вертикали - количество интервалов; по горизонтали - их длительность в мс. Сплошная горизонтальная линия - среднее количество интервалов; штриховая - уровень 2s .
На (В) представлен нейрон, у которого в течение 3 ч после введения резерпина частота фонового разряда существенно не менялись, а его структура изменялась недостаточно отчетливо. Резистентные к резерпину нейроны преимущественно залегали на уровне 4-го слоя колонки, т.е. они в большинстве своем были входными для первичной специфической афферентации. Средний межимпульсный интервал у резистентных к резерпину нейронов (N=10) в контроле был равен 15,4± 5,0 мс, а через 2-3 часа после введения резерпина - 20,2± 6,0 мс, что по t- критерию недостоверно отличается от контроля (P<0,05).
Оценка достоверности различия выборочных распределений межимпульсных интервалов у представленных трех типов нейронов по критерию Фишера показала достоверную (P<0,05) трансформацию под влиянием резерпина структуры их фонового импульсного разряда (F=1,76; 2,88; 1,46, соответственно, для А, Б и В при Fst=1,45).
С помощью метода пошагового определения плотности активных в фоне нейронов в отвесных проходках через кору мозга исследованы в трех острых опытах 9 микроэлектродных треков, которые проходили, как показала пороговая идентификация, в пределах бочонковых колонок представительства средних вибрисс рядов В и С. В первом опыте в контроле наблюдалось 29, во втором опыте - 27 и в третьем - 25 фоновых нейронов, что составило в контроле в среднем на один трек 9 нейронов. Более плотно активные в фоне нейроны располагались на глубине 500-1500 мкм - 70% от общего количества. Суммарно по трем опытам в контроле было зарегистрировано на уровне: 2-3 слоев - 18 нейронов, 4 слоя -26 нейронов, 5 слоя - 23 нейрона, 6 слоя - 14 нейронов. В исходном состоянии на треках встречалось от 4 до 18 фоновоактивных нейронов. Как и в серии опытов с электрической стимуляцией ядер шва, треки с высокой плотностью нейронов были разделены относительно пустыми" треками. В шести треках с высокой плотностью элементов (7 - 18) зарегистрировано 67 фоновых нейронов (83%), а в трех относительно "пустых" треках (4 - 5 нейронов) - 14 нейронов (17%).
Пространственно регулярный профиль распределения фоновоактивных нейронов в ПМСБ CI коры мозга крыс сохранялся и на фоне резерпина. Спустя 2-3 часа после введения резерпина в тех же треках в первом опыте зарегистрирована активность 21 нейрона, во втором опыте - 22 нейронов и в третьем - 21 нейрона, что составило в среднем около 7 нейронов на трек. Из них зарегистрировано на уровне: 2-3 слоев - 10 нейронов, 4 слоя - 23 нейрона, 5 слоя - 21 нейрон, 6 слоя - 10 нейронов. В шести треках с высокой плотностью элементов при этом зарегистрировано - 52 нейрона (81%), а в трех "пустых" треках - 12 нейронов (19%). Из данных послойного анализа следует, что резерпин наиболее существенно уменьшает количество фоновоактивных нейронов в бочонковых колонках на уровне 2-3 слоев (на 44%) и 6 слоя (на 29%). Плотные и относительно "пустые" треки в последействии резерпина содержали, соответственно, на 27% и на 25% меньше активных в фоне нейронов.