Гены идентичности цветковых меристем
Мутации в локусе арабидопсиса TERMINAL FLOWER1 (TFL I) приводят к фенотипу, противоположному таковому для мутаций по генам идентичности цветковой меристемы. Апикальная и латеральная меристемы соцветия у них заменяются цветковыми меристемами. Кроме того, как тоже упоминалось ранее, мутанты tfll цветут раньше растений дикого типа. В отличие от растений арабидопсиса дикого типа, имеющих недетерминированное соцветие с множеством цветков и вторичными и третичными соцветиями, мутанты tfll имеют детерминированное первичное соцветие всего лишь с несколькими цветками и не образуют вторичных соцветий. Соцветие заканчивается аномальным цветком, состоящим из двух-трех неполных цветков. Похоже, вторичные меристемы соцветия у мутантов tfll заменяются меристемами цветка, а первичная генеративная меристема превращается в 2—3 смежные цветковые меристемы. Можно предположить, что ген TFL1 необходим для поддержания функциональности, или компетентности, цветковой меристемы, хотя, по-видимому, он активен и в вегетативной фазе развития. TFL1 также играет роль в негативной регуляции экспрессии генов LFY и API в апикальных и латеральных меристемах, хотя такая регуляция может осуществляться и опосредованно.
Гомологом гена TFL1 у львиного зева является ген CENTRO-RADIALIS (CEN). В норме соцветия львиного зева, как и арабидопсиса, являются недетерминированными: цветки образуются в латеральных положениях, и никогда не формируется терминальный, или апикальный, цветок, который был бы последним цветком в соцветии. При этом гены FLO, LFY, SQUA и API экспрессируются в латеральных меристемах, но не в апикальной части соцветия. Однако в апексах меристемы мутантов сеп, как и tfll, образуются терминальные цветки. Как и ожидалось, вышеперечисленные гены экспрессируются в апексах меристем этих мутантов. Более того, условия окружающей среды, которые усиливают фенотип Ify, например короткий день, ослабляют фенотип tfll. Таким образом, гены CEN и TFL1, по-видимому, являются антагонистами генов, индуцирующих развитие цветка, и, возможно, могут предотвращать их экспрессию в апексе соцветия. На основе анализа фенотипов двойных мутантов предполагают, что TFL1 является антагонистом генов LFY, API и АР2. Мутации Ify частично супрессируют tfll, двойные мутанты Ify apl или Ify ap2 характеризуются более выраженным супрессирующим эффектом на фенотип tfll. Терминальный цветок мутантов сеп обладает радиальной симметрией: все его лепестки напоминают вентральные лепестки цветков дикого типа. Количество органов цветка сеп и их филотаксис (расположение} очень вариабельны
Глава 2. Молекулярная характеристика генов, контролирующих идентичность цветковой меристемы
2.1 Молекулярная характеристика генов FLO (львиный зев) и LFI (арабидопсис)
Ген львиного зева FLO был клонирован одним из первых генов, влияющих на развитие цветка. Для его клонирования использовали мутант По-613, образующий генеративные соцветия вместо цветков. Однако гомозиготы по flo-613 изредка формировали нормальные цветки. Из семян этих цветков развивались растения дикого типа. Значит, мутация flo-613 генетически нестабильна и иногда ревертировала к аллели дикого типа. Реверсия коррелировала с эксцизией транспозона ТатЗ и, следовательно, мутантный фенотип был обусловлен инсерцией этого мобильного элемента в ген FLO. Таким образом, ген FLO был клонирован с использованием последовательности транспозона ТатЗ в качестве пробы. У всех ревертантов к дикому типу при использовании клонированного фрагмента гена FLO как пробы для гибридизации появлялся фрагмент длиной 4т. п. о. Такой же длиной обладал гибридизовавшийся с FLO фрагмент исходного растения дикого типа, использованного для транспозонового мутагенеза. При этом у некоторых ревертантов сохранялся фрагмент длиной 7.5 т. п. о., такой же, как и у мутанта flo-613. Вероятно, среди ревертантов были как гетерозиготные, так и гомозиготные растения.
Однако при гибридизации ДНК мутантных растений среди них тоже были выявлены гетерозиготы, у которых присутствовали оба фрагмента. Так, при секвенировании сайта эксцизии ТатЗ из гетерозиготы с фенотипом По обнаружили инсерцию 8 пар оснований. Таким образом, неточная эксцизия ТатЗ приводила к изменению рамки считывания, и мутантный фенотип сохранялся.
В сайте эксцизии у ревертанта отмечены замены двух пар оснований, и не обнаружены инсерции или делеции нуклеотидов. В потомстве от самоопыления всех гетерозиготных ревертантов имеются растения дикого типа и мутанты в соотношении 3:3. Потомство большинства гомозиготных ревертантов состояло только из растений дикого типа.
Мутация flo-613, как и многие другие гомеозисные мутации львиного зева, использованные для клонирования генов развития цветка и о которых пойдет речь в этой главе, была получена в массивном эксперименте по транспозоновому мутагенезу. Растения львиного зева с высокоактивными мобильными элементами Тат (от Transposone of Antirrhinum majus) выращивали при температуре 15 °С. Именно при этой температуре для мобильных элементов группы Тат наблюдается наибольшая частота транспозиций. Многие из 26000 потомков Ml этих растений, полученных при самоопылении, содержали рецессивные мутации в гетерозиготном состоянии. Такие рецессивные мутации можно было выявить в следующем поколении М2 после самоопыления растений Ml. И в самом деле, среди 80000 растений М2 было обнаружено более 15 независимых гомеозисных мутаций, затрагивающих развитие цветка (рис.2).
Ген арабидопсиса LFY был клонирован по гомологии с геном львиного зева FLO. В отличие от многих других гомеозисных генов, затрагивающих развитие цветка, FLO и LFY, похоже, уникальные гены и не являются членами генного семейства. Кодируемые ими белки были на 70 % гомологичны друг другу и не обладали значительной гомологией ни с одним из известных на момент их клонирования белков. Наличие богатого пролином домена на N-конце и кислой области в центральной части белка свидетельствуют о том, что, вероятно, они могут служить активаторами транскрипции. Подтверждается это и тем, что белок Lfy локализован в основном в ядре. Характер их экспрессии очень похож. РНК FLO и LFY начинает накапливаться в закладывающихся зачатках цветка на границе цветочной меристемы и не обнаруживается в генеративной меристеме. В момент приобретения цветочными меристемами морфологических различий (стадии 1 и 2) экспрессия усиливается в равной степени по всему молодому зачатку цветка. На стадиях 3 и 4, когда индуцируется экспрессия генов идентичности органов, РНК FLO и LFY уже практически не обнаруживается в центре начинающего формироваться цветка. Из известных генов они первыми начинают экспрессироваться в клетках, из которых затем сформируется цветок.
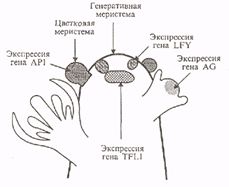
Рис.2. Схематическое изображение генеративного побега арабидопсиса и начальные этапы экспрессии генов, регулирующие развитие цветка.
Однако в характере экспрессии FLO и LFY есть одно важное отличие, помогающее понять, как развиваются меристемы. Различие является следствием того, что у цветка львиного зева есть прицветники, а у цветка арабидопсиса их нет. Ген FLO экспрессиру-ется как в примордиях прицветников, так и в цветковых меристемах. В то же время для гена LFY не наблюдается экспрессия в клетках, окружающих цветковые меристемы, и, вероятно, соответствующих клеткам львиного зева, из которых образуются прицветники. У мутантов Ify на месте цветков дикого типа без прицветников образуются генеративные побеги или «цветки» с околоплодными листьями, которые можно отождествлять с прицветниками. Значит, продукт LFY дикого типа подавляет инициацию или образование прицветников. Предполагают, что ген LFY влияет на судьбу группы клеток на периферии цветочной меристемы: в цветках дикого типа все или большинство этих клеток формируют цветочную меристему, тогда как у мутантов Ify эти клетки дают начало меристеме и примордию прицветника. В этом отношении мутант Ify похож на дикий тип растений таких видов, как Antirrhinum majus, у которых из одного зачатка образуются меристема цветка и прицветник. Считается, что наличие прицветников - эволюционно более древний признак.