Зрительные пигменты
Если мы проецируем на белый экран два пучка света различных цветов одновременно . мы видим только один цвет, более или менее отличный от обоих этих цветов. Мы сможем лучше понять выдающийся факт того, что мы способны воспринимать все оттенки в составе внешнего света путем смеси трех основных цветов, если мы сравним глаз сухом . В случае звука . мы слышим более длинные волны как низкие тона, а короткие волны - как высокие и пронзительные, кроме этого ухо способно улавливать одновременно много звуковых волн, т.е. много нот. Однако они β данном случае не сливаются в один сложный аккорд, подобно тому, как различные цвета . сливаются в один сложный цвет. Глаз не может показать разницу, если мы заменяем оранжевый цвет на красный или желтый; но если мы слышим ноты до и ми, звучащие одновременно, нам подобное звучание не кажется нотой ре. Если бы ухо воспринимало музыкальные тона подобно тому, как глаз воспринимает цвета, каждый аккорд мог бы быть представлен комбинацией трех постоянных нот, одной очень низкой, одной очень высокой и одной промежуточной, вызывая все возможные музыкальные эффекты только путем изменений относительной громкости этих трех нот . Однако мы способны видеть плавный переход цветов одного в другой через бесконечное множество оттенков и градаций . То, каким образом мы воспринимаем каждый из цветов . зависит е основном от строения нашей нервной системы. Надо признаться, в настоящее время ни у человека, ни у четвероногих не описана анатомическая база для подтверждения теории восприятия цвета.
Эти точные и дальновидные предсказания были подтверждены серией различных наблюдений. При помощи спектрофотометрии Вальд, Браук, МакНикол и Дартнэл с коллегами показали наличие в сетчатке человека трех типов колбочек с различными пигментами. Также Бейлор с коллегами сумели отвести токи от колбочек обезьян и человека. Было обнаружено, что три популяции колбочек имеют различные, но перекрывающиеся диапазоны чувствительности к голубой, зеленой и красной части спектра. Оптимальные длины волн для возбуждения электрических сигналов в точности совпали с пиками поглощения света зрительными пигментами, установленными при помощи спектрофотометрии и при психофизических экспериментах по измерению чувствительности глаза к цветовому спектру. В конечном итоге Натаисом были клонированы и секвенированы гены, кодирующие пигмент опсин в трех типах колбочек, чувствительных к красному, зеленому и голубому спектру.
Каким же образом молекулы различных зрительных пигментов способны предпочтительно улавливать свет определенной длины волны? Оказывается, родопсин - зрительный пигмент палочек и все три зрительных пигмента колбочек содержат в своем составе один и тот же хромофор, 11-цис-ретиналь. Однако аминокислотные последовательности белковой части пигмента отличаются друг от друга. Различиями всего в нескольких аминокислотах и объясняется различная их чувствительность к спектру.
Цветовая слепота
Хотя один тип фоторецептора не способен сам по себе воспринимать цвет, три типа колбочек, как показано на рис. 4, уже способны.
В принципе, двух типов колбочек с различными пигментами было бы достаточно для распознавания цвета, однако в этом случае ряд комбинаций длин волн воспринимался бы одинаково. Подобная ситуация возникает в случае, когда человек страдает цветовой слепотой. У таких людей, как было показано Натансом, имеются генетические дефекты, приводящие к отсутствию одного из пигментов. С высоты нынешнего состояния науки мы не можем не поразиться тому, как красиво молекулярные механизмы подтверждают блистательные и на удивление точные размышления Янга и Гельмгольца.
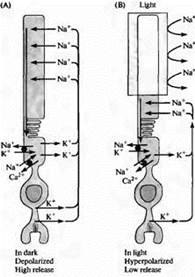
Рис. 4. "Темновой" ток в палочке. (А) В темноте ионы натрия проходят через катионные каналы в наружном сегменте палочек, вызывая деполяризацию; ионы кальция также способны проходить через зти катионные каналы. Петля тока проходит через перешеек палочки за счет того, что калиевый ток течет в направлении наружу во внутреннем сегменте мембраны. (В) Когда происходит освещение наружного сегмента, каналы закрываются из-за уменьшения внутриклеточной концентрации цГМФ, и палочка гиперполяризуется. Гиперполяризация проводит к уменьшению высвобождения медиатора. Концентрации натрия, калия и кальция поддерживаются внутри палочки специальными насосами и ионообменниками, расположенными в области внутреннего сегмента (черные кружки). Транпортеры кальция также находятся и в наружном сегменте.
Их идея о том, что основные атрибуты цветного зрения и цветовой слепоты должны быть обнаружены в самих фоторецепторах, были подтверждены при помощи прямых физиологических измерений, а также при изучении различий в строение пигментов на генетическом и белковом уровнях ный потенциал составляет порядка - 40 мВ, что далеко от равновесного калиевого потенциала ЕК, составляющего - 80 мВ.
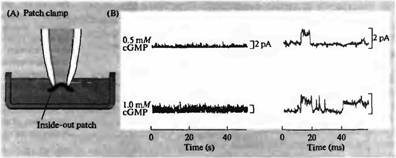
Рис. 5. Роль цГМФ в открытии натриевых каналов наружного сегмента палочек. Проводилась регистрация активности одиночных каналов при помощи inside-out patch clamp, находящихся в растворе с разной концентрацией цГМФ. Открытие канала приводит к отклонению тока вверх. Частота открытия каналов была очень низка в контроле, добавление цГМФ приводило к увеличению частоты открытия одиночных каналов, которая прямо зависела от концентрации цГМФ
Входящий ток в темноте переносится в основном ионами натрия, следующими по электрохимическому градиенту через катионные каналы наружного сегмента. Гиперполяризация фоторецептора при действии света обусловлена закрытием этих каналов, что приводит к сдвигу мембранного потенциала в сторону ЕК.
Свойства каналов фоторецептора
Катионные каналы наружного сегмента при физиологических условиях имеют соотношение в проводимости ионов кальция/натрия/калия как 12,5: 1,0: 0,7 и проводимость одиночного канала около 0,1 пСм21). Из-за того, что концентрация натрия гораздо выше, чем концентрация кальция, около 85% входящего тока приходится на ионы натрия. Движущая сила ионов калия направлена наружу клетки. При прохождении канала ионами кальция, эти ионы прочно связываются со стенкой поры и мешают, таким образом, проведению других ионов. Именно из-за этого удаление кальция из внеклеточной среды приводит к более легкому прохождению ионов калия и натрия через каналы, проводимость которых возрастает до 25 пСм.
Фесенко, Яу, Бейлор, Страйер с коллегами показали, что циклическая ГМФ играет роль внутреннего переносчика сигнала от диска к поверхности мембраны. Как показано на рис.4, высокая концентрация цГМФ в цитоплазме поддерживает катионные каналы в открытом состоянии. При уменьшении концентрации цГМФ с внутренней поверхности мембраны открытие катионных каналов становится редким событием. Таким образом, мембранный потенциал фоторецепторов является отражением концентрации цГМФ в цитоплазме: чем выше концентрация цГМФ, тем больше деполяризация клетки. Концентрация же цГМФ, в свою очередь, зависит от интенсивности падающего света. Повышение интенсивности света приводит к уменьшению концентрации цГМФ и уменьшает долю открытых каналов. В отсутствии цГМФ почти все каналы закрыты, и сопротивление мембраны наружного сегмента приближается к значению сопротивления липидного бислоя.