Зрительные пигменты
Молекулярная структура цГМФ-управляемых каналов
Была выделена кДНК для ионных каналов наружного сегмента палочек и определены аминокислотные последовательности субъединиц данных каналов в сетчатке человека, быка, мыши и курицы. Было обнаружено значительное сходство ДНК для этих и других цГМФ-управляемых ионных каналов - например, найденных в обонятельной
Передача сигнала в фоторецепторах
Каким образом фотоизомеризация родопсина приводит к изменению мембранного потенциала? На протяжении многих лет было понятно, что необходим какой-то внутренний посредник для возбуждения электрических сигналов в палочках и колбочках. Одной из причин подозревать, что информация о поглощении фотонов в области наружного сегмента палочки передается при помощи переносчика, был тот факт, что сам пигмент родопсин находится внутри диска, а сигнал распространяется через цитоплазму до внешней мембраны. Второй причиной было значительное усиление ответа. Бейлор с коллегами, изучая фоторецепторы черепахи, показал, что уменьшение мембранной проводимости и регистрируемых электрических сигналов возникает уже при поглощении всего одного фотона и активации одной из 108 пигментных молекул.
Последовательность событий, при которых активировавшаяся молекула фотопигмента изменяет мембранный потенциал, была выяснена при изучении наружных сегментов палочек и колбочек, используя методы двухэлектродной фиксации потенциала и методов молекулярной биологии. Схема передачи сигнала от поглощения фотона света до электрического сигнала показана на рис. 3.
В темноте в наружных сегментах палочек и колбочек протекает постоянный входящий "темновой" ток. В результате их мембран системе. Мембранные участки этих каналов проявляют структурные сходства с другими катион-селективными каналами, особенно в области S4 и в области, формирующей ионную пору. Ионные каналы фоторецептора являются тетрамерами, составленными, по крайней мере, из 2 различных белковых субъединиц с молекулярной массой 63 и 240 кДа соответственно.
Внутриклеточные места связывания нуклеотидов расположены около карбоксильного конца субъединиц. Экспрессия этих субъединиц в ооцитах приводит к формированию катионных каналов, имеющих свойства подобные каналам, расположенным в наружных сегментах палочек: они активируются цГМФ и имеют ожидаемые соотношения проводимости и проницаемости.
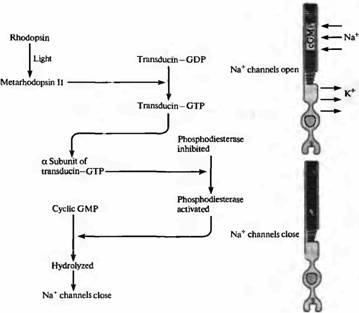
Рис. 6. Механизм активации G-белка при активации молекулы фотопигмента. G-белок трансдуцин связывает ГТФ в присутствии метародопсина II, что приводит к активации фосфодиэстеразы, которая, в свою очередь, гидролизует цГМФ. При падении концентрации цГМФ закрываются натриевые каналы.
Метаболический каскад циклического ГМФ
Цепочка событий, приводящих к уменьшению концентрации цГМФ и к последующему закрытию ионных каналов, показана на рис.5. Уменьшение внутриклеточной концентрации цГМФ вызывается светом, приводящим к образованию метародопсина II, промежуточного продукта в распаде фотопигмента. Метародопсин II воздействует, в свою очередь, на G-белок трансдуцин, который состоит из 3 полипептидных цепей·
Взаимодействие метародопсина II и трансдуцина приводит к замене связанной с G-белком молекулы ГДФ на ГТФ. Это активирует ?субъединицу G-белка, которая отделяется от субъединиц и, в свою очередь, активирует расположенную в примембранной области фосфодиэстеразу: фермент, который гидролизует цГМФ. Концентрация цГМФ падает, становится меньше открытых ионных каналов, палочка гиперполяризуется. Каскад прерывается при фосфорилировании С-конца активного метародопсина II. Ключевая роль цГМФ в регулировании состоянии катионных каналов была подтверждена биохимическими экспериментами. Освещение фоторецепторов может вызвать 20% падение уровня цГМФ в клетке.
Рецепторы позвоночных, деполяризующиеся при действии света
Интересным исключением из механизма фоторецепции, приведенного выше, являются некоторые рецепторы позвоночных. У ящериц есть третий глаз, расположенный на макушке. В нем находятся маленькие "колбочки", способные воспринимать изображение, сходное с воспринимаемым основными (боковыми) глазами. Эти фоторецепторы, однако, примечательны тем, что деполяризуются при освещении. Каналы, управляемые нуклеотидами, здесь имеют сходное строение и функционирование с фоторецепторами других позвоночных, за одним исключением: активация фоторецептора и G-белка приводит к увеличению концентрации цГМФ. В результате этого каналы наружного сегмента открываются, и катионы направляются внутрь клетки, формируя "световой" ток. Это происходит благодаря угнетению активности фосфодиэстеразы в темноте. В результате в глазу ящерицы происходит следующая последовательность событий: свет - [цГМФ] - ткрытие катионных каналов наружного сегмента - деполяризация.
Усиление сигнала в каскаде цГМФ
Двухэтапный каскад цГМФ обеспечивает значительное усиление исходного сигнала, чем и объясняется исключительная чувствительность палочек к свету. Во-первых, одна молекула активного метародопсина II катализирует присоединение многих молекул ГТФ вместо ГДФ и, таким образом, освобождает сотни субъединиц G-белка. Во-вторых, каждая субъединица активирует одну молекулу фосфодиэстеразы в диске, которая способна расщепить огромное количество молекул цГМФ, находящихся в цитоплазме, и таким образом привести к закрытию большого количества каналов.
Сигналы в ответ на одиночные кванты света
Данные о том, что одиночные кванты света могут вызывать воспринимаемое ощущение света, вызвали большое количество вопросов. Насколько велик этот единичный ответ? Каким образом этот сигнал выделяется из уровня шума? И каким образом такая информация достоверно передается из сетчатки в высшие зрительные центры? Чтобы измерить сигналы в ответ на одиночные кванты света, Бейлор с коллегами регистрировали токи от отдельных палочек в сетчатках жабы, обезьяны и человека. Эти опыты представляют собой уникальный пример эксперимента, в котором показано, как такой сложный процесс, как восприятие слабых вспышек света, может коррелировать с изменениями, происходящими на уровне отдельных молекул.
Процедура по изоляции фрагмента сетчатки от животного либо трупного материала должна для этих экспериментов производиться в темноте. Для измерения тока наружный сегмент палочки засасывается в тонкую пипетку. Как и ожидалось, эти эксперименты показывают, что в темноте ток постоянно течет внутрь наружного сегмента. Вспышки света приводят к закрытию каналов в наружном сегменте, вызывая уменьшение "темнового" тока. Амплитуда токов невелика и пропорциональна числу поглощенных квантов. Иногда вспышка вызывает одиночный ответ, иногда - двойной, а иногда - вообще никакой реакции.
В палочках обезьяны уменьшение тока в ответ на поглощение одного фотона, составляет около 0,5 пА. Это соответствует закрытию порядка 300 каналов, т.е. от 3 до 5% всех открытых в темноте каналов. Это достигается благодаря значительному усилению сигнала в метаболическом каскаде цГМФ. Более того, из-за крайней стабильности зрительных пигментов, упомянутой ранее, случайная изомеризация и ложное закрытие каналов - события очень редкие. Это приводит к тому, что эффекты отдельных квантов света выделяются на фоне очень низкого постоянного шума. Было показано, что электрическая связь посредством щелевых контактов между фоторецепторами обеспечивает дополнительный сглаживающий эффект, который уменьшает фоновый шум и улучшает отношение сигнал/шум ответов палочек на одиночные кванты.